 2006, Vol. 42
2006, Vol. 42文章信息
- 张清海, 叶功富, 林益明.
- Zhang Qinghai, Ye Gongfu, Lin Yiming.
- 海岸退化沙地木麻黄人工林能量的研究
- Studies on Energy of Casuarina equisetifolia Plantation on Degraded Coastal Sand
- 林业科学, 2006, 42(8): 1-7.
- Scientia Silvae Sinicae, 2006, 42(8): 1-7.
-
文章历史
- 收稿日期:2005-01-26
-
作者相关文章



2. 福建林业科学研究院 福州 350012;
3. 厦门大学生命科学学院 厦门 361005
2. Fujian Academy of Forestry Fuzhou 350012;
3. School of Life Sciences, Xiamen University Xiamen 361005
木麻黄(Casuarina equisetifolia)自20世纪50年代引种中国已经有50多年的历史,是我国东南沿海地区主要造林树种,特别是在沿海沙地上,已成为当家树种。沿海防护林是沿海岸主要的生态屏障,以木麻黄为主的人工生态系统,在防风护沙、改善沿海生态环境和促进经济建设等方面起到难以替代的作用。滨海沙地属极度退化立地,适生树种少,构建可持续经营的人工生态系统难度大。Gupta(1972)、Jordan(1971)研究认为,应用能量的概念研究植物群落比单纯用干物质测定更能反映出群落对自然资源(特别是太阳能)的利用情况。本文从能量生态学角度,研究木麻黄人工林的能量特征,深入了解沿海基干防护林带生态系统的结构与功能,为沿海地区退化立地的生态恢复、现有防护林的更新改造及实现可持续经营提供科学依据。
1 试验地概况试验地设在福建省沿海中部惠安县崇武赤湖防护林场(118°55′E,23°45′N)。属南亚热带海洋性季风气候,年均气温19.8 ℃,最高气温37 ℃,最低气温2.2 ℃,全年无霜期320 d,年均降水1 029 mm,年均蒸发量2 000 mm,蒸发量大于降雨量,干湿季明显,夏季(7—9月)多台风和暴雨,秋冬东北风强劲,8级以上大风105 d,年均风速7.0 m· s-1,干湿季明显,干旱频繁发生。土壤为均一性风积沙土,沙土层厚度80~100 cm。
赤湖林场,1989年造,密度2 m×2 m(2 500株·hm-2),现密度为1 648株·hm -2,平均胸径10.77 cm,平均树高12.97 m,郁闭度大于0.90。林地内凋落物厚度3 ~5 cm,分解良好。林下灌木、草本稀少。“九五"期间建立了固定生态定位观察点,试验地不受人为干扰和破坏。
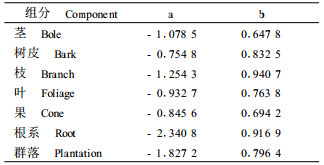
2 试验方法 2.1 生物量的测定在试验地中设立25 m×25 m样地3块。对样地内的林木进行每木检测,根据每木测定的胸径和树高值选取平均木,按2 m区分段进行树干解析,枝叶部分采用分层收获法,地下部分采用分层壕沟法,现场称取样木各组分的鲜质量,并取样回实验室在80 ℃下烘干,另取小样在1 05 ℃下烘干,求含水率。根据各组分的含水率,求各组分的干物质量,根据标准木树干解析推算各林分的生物量。初级净生产力根据前人的经验公式lnW=a+bln(D2H)计算:群落能量的净生产力根据年初对群落中胸径和树高测定值,系数a和b引用前人的研究成果(叶功富等,1996a),见表 1。
|
|
凋落物的收集采用收集框法,每个样地林下随机设置9个收集筐,为长1 m, 宽1 m, 深15 c m,孔径0.5 mm的玻璃纤维网,筐底离地面20 cm,每15 d收集1次,及时烘干,每月收集2次并进行汇总,分出叶(小枝)、枝、果和花,分别测定鲜质量,在80 ℃下烘干,各组分另取少量在105 ℃下烘干到恒重,量少的组分则在80 ℃下烘干至恒重,求算每月凋落物的总量,对每月的资料汇总得全年总量(林益明等,2001;叶功富等,1996b)。
2.3 热值的测定所有采集样品在80 ℃烘干,磨粉处理后过6号筛贮存备用,然后用热量计测定热值,仪器采用长沙仪器厂生产的GR-3500型微电脑氧弹式热量计。样品热值以干质量热值(每克干物质在完全燃烧条件下所释放的总热量,简称GCV)和去灰分热值(AFCV)表示。空调控制测定环境20 ℃,每样品重复多次,误差控制在±0.20 kJ·g-1,每次测试时对仪器用苯甲酸标定(祖元刚,1990;林益明等,2001)。
灰分含量测定采用干灰化法,样品在马福炉中550 ℃下灰化5 h,然后测定灰分含量。去灰分热值=干质量热值/(1-灰分含量)。去灰分热值可以除去不同环境下灰分含量不同的差异对热值的影响,更能够反映植物体各组分热值情况(林益明等,2001),特别是叶,更能体现其对太阳能的利用率。
能量现存量根据各组分的平均热值与各组分的生物量现存量相乘累加而得。
3 结果与分析 3.1 生物量生物量是指群落中一定时间内单位面积上的物质数量,通常是指活的植物体及其上枯死的植物体(林鹏等,1999)。
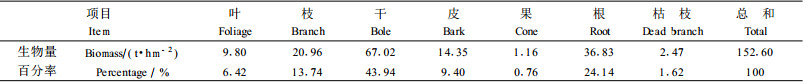
从表 2可知:林分中各组分现存总生物量为152.60 t·hm-2,其中干的生物量最大为67.02 t·hm-2,占整个林分总量的43.94%,果实的生物量最小为1.16 t·hm -2,占整个林分总量的0.76%,各组分的具体情况为:干(43.94%)>根(24.14%)>枝(13.74%)>皮(9.40%)>叶(6.42 %)>枯枝(1.62%)>果(0.76%)。整个林分生物量与南亚热带其他林分相比,比鹤山17年生马占相思(Acacia mangium)林124.23 t·hm-2 (彭少麟等,1998)、惠安赤湖林场15年生木麻黄123.76 t·hm-2 (叶功富等,199 6a)、印度喜马拉雅山中部贫瘠砾石地上15年生的印度黄檀(Dalbergia sissoo)与桃金娘(Rhodomyrtus tomentosa)混交林118.10 t·hm-2(Lodhiyal et al., 2003)、香港的桃金娘灌丛36.47 t·hm-2(管东生,1998)均高。但比鼎湖山的锥栗(Castanopsis chinensis)、黄果厚壳桂(Cryptocarya concinna)和荷木(Schima superba)组成的天然林292.98 t·hm-2(温达志等,1997)、惠安20年生木麻黄人工林194.31 t·hm-2(吴寿德等,1996)、中亚热带20年生檫木(Sassafras tzumu)林586.93 t·hm-2(柳江等,2002)均低。地上部分与地下部分生物量比为3.14:1,根占总生物量的24.14%,具有较高的含量,比鼎湖山格木(Erythrophleum fordii)天然林14.41%(蚁伟民等,2000)和宝天曼自然保护区45年生栓皮栎(Quercus variabilis)林22.29%(刘玉萃等,1998)高。McConnaughay等(1999)和Odum (2002)研究认为植物总是尽量把生物量分布于受限最大的器官上,当土壤缺少养分时根系的生物量比重增大。滨海沙地养分含量低,是木麻黄生长的一个主要限制因素,木麻黄必须具有高度发达的根系,来维持其快速生长对养分的需求。木麻黄根系分布较浅,主根不明显但侧根发达,有较高生物量,有着很好的固沙作用。总之木麻黄在极度退化的沙地上具有较高的现存生物量,体现出对滨海沙地有很好的适应能力。
|
|
净初级生产力(net primary productivity)是指林分中除去其生理活动代谢和动物啃食之外,单位面积单位时间内林分所固定的生物量的总量。木麻黄林各组分净初级生产力见表 3。
|
|

木麻黄人工林年净生产力为10.17 t·hm-2a-1,比惠安10年生木麻黄7.97 t ·hm-2a-1(叶功富等,1996b)、南亚热带香港的桃金娘灌丛10.10 t·hm-2 a-1 (管东生,1998)、夏威夷的红椿(Toona cilata)林8.40 t·hm -2 a-1 (Ares et al., 2000)均高,与武夷山黄山松(Pinus taiwanensis)林10.18 t·hm-2a-1相近(林鹏等,1999),比印度喜马拉雅山中部贫瘠砾石地上印度黄檀林14.18 t·hm-2a-1 (Lodhiyal et al., 2003)低,可见木麻黄是具有较高生产力水平的沙地速生树种,其在贫瘠沙地的适应性很好。树干年净生产力最大5.72 t·hm-2a-1,占总量的56.24%,果年净生产力最小0.12 t·hm-2a-1,占总量的1.21%,各组分的具体情况如下:干(56.24%)>根(14.31%)>枝(10.30%)>叶(9.82%)>皮(8.12%)>果(1.21%)。
3.3 凋落物归还量凋落物是生命系统向环境归还物质和能量的主要形式,同时也是生命系统与环境之间进行物质、能量、信息交换的主要形式。
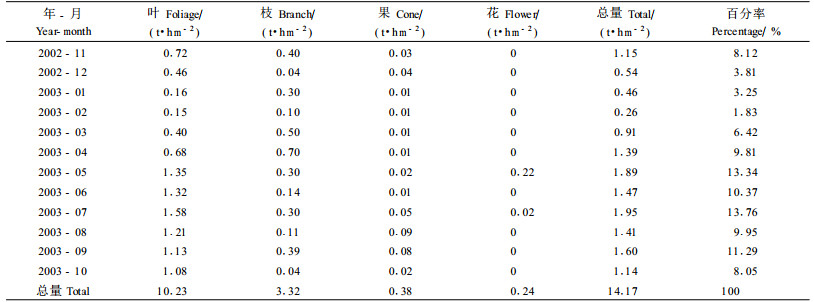
滨海木麻黄人工林具有较高的凋落物归还量14.17 t·hm-2a-1,比惠安15年生木麻黄2.17 t·hm-2a-1(叶功富等,1996b)、印度喜马拉雅山中部贫瘠砾石地上15年生的印度黄檀林4.37 t·hm-2a-1 (Lodhiyal et al., 2003)、南亚热带鹤山的马占相思林4.90 t·hm-2a-1 (彭少麟,2003)均高。从表 4可知,5—9月归还的凋落量占总量的58.65%,与Srivastava(1995)研究结果(5月份凋落物归还量最大)相似。这个期间是台风多发的季节,雨量较高,木麻黄生长和代谢旺盛;同时台风也造成了大量的凋落物。年初凋落物量最小,这与气温回升不高,又属旱季,木麻黄生长较慢及代谢慢有关。凋落物主成分是叶,年归还量为10.23 t·hm-2a -1,占总归还量的72.19%,比Strivastava(1995)研究结果87%~95%低;其次是枝条,年归还量3.32 t·hm-2a-1,占总凋落物量的23.43%;再次是果0.38 t·hm-2a-1,占总量的2.68%;其余部分0.24 t·hm-2a-1,占总量的1.69%。
|
|
热值是单位质量的物质在常温常压强下, 完全燃烧所释放出来的热量。植物各组分的热值是各组分干物质单位质量在常温常压下完全燃烧所释放的热量,通常称干质量热值,以下文中出现热值即指干质量热值。热值是计算群落能量的基础,在生物量和热值的基础上可以求算群落的现存能量。植物体热值是衡量植物光能利用和生产力的指标之一(林鹏等, 1999)。
木麻黄人工林各组分的热值、灰分含量、去灰分热值见表 5,叶热值的空间分布情况为:上层叶最高20.53 kJ·g-1,其次为中层叶,20.07 kJ·g-1,下层叶最小,19.44 kJ·g-1,总体加权平均值为20.01 kJ·g-1。叶的热值从上往下减小主要是林分中太阳辐射从上往下逐渐减少造成的。枝条与叶呈相反的趋势:上层枝最小,19.26 kJ·g-1,其次中层枝,下层枝最大,总体平均热值(林分总能量与林分总生物量的比值)为19.70 kJ·g-1,比施肥林地银柳(Salix discolor)林的19.22 kJ·g-1、Salix petiolaris林的19.59 kJ·g-1、蒿柳(Salix viminalis)林的19.21 kJ·g-1 (Labrecque et al., 1997)均高。各组分热值顺序为:果(20.23 kJ·g-1)>叶(20.01 kJ·g-1) >枯枝(19.83 kJ·g-1)>根(19.56 kJ·g-1) >皮(19.51 kJ·g-1)>枝(19.41 kJ·g-1)>干(19.29 kJ·g-1)。
|
|
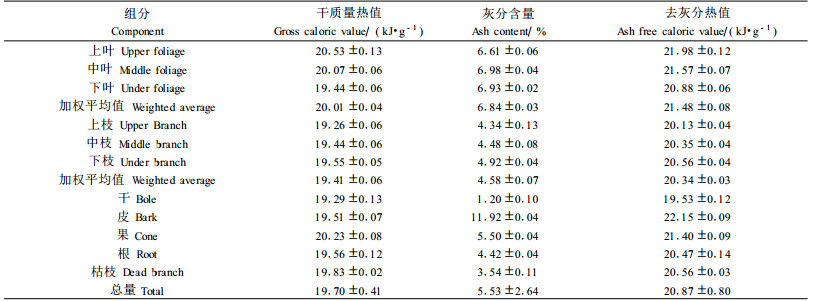
灰分含量是植物体对元素聚集度的体现,特别是金属元素。皮的灰分含量最高,11.92%,干的灰分含量最小,1.20%,总体平均为5.23%, 各组分的大小顺序为:皮(11.92%)>叶(6.84%)>果(5.50%)>枝(4.58%)>根(4.42%)>枯枝(3.54%)>干(1.20%)。
去灰分热值是去除由于元素的聚集不同造成的热值差异,更能体现各组分的热值。其中上层叶的去灰分热值最大,21.98 kJ·g-1,干最小,19.53 kJ·g-1。各组分的平均去灰分热值为:皮(22.15 kJ·g-1)>小枝(21.48 kJ·g-1) >果(21.40 kJ·g-1) >枯枝(20.56 kJ·g-1)>根(20.47 kJ·g-1)>枝(20.3 4 kJ·g-1) >干(19.53 kJ·g-1)。
用SPSS10.0对热值与灰分含量分析,结果表明:小枝热值与灰分含量线性负相关,相关系数r为-0.71,P < 0.05;枝热值与灰分含量呈线性正相关,r为0.74,P < 0.05。两者存在差异是由不同组分具有不同的结构和元素含量造成的。
1年内凋落物各组分热值的测定结果见表 6。小枝平均热值为21.11 kJ·g-1,与和溪亚热带雨林凋落物叶21.33 kJ·g-1 (林鹏等,1995)相近,枝条19.89 kJ·g -1,果19.91 kJ·g-1,花20.96 kJ·g-1,均低于和溪亚热带雨林(林鹏等,1995)。
|
|

群落内能量的组合特征是群落各组分在长期进化与演替过程中形成的, 是群落各组分之间及其与环境之间长期相互作用、相互适应的结果, 是能量长期积累的一种表现形式, 是衡量不同生态系统结构与功能基础的最佳可比指标(孙雪峰等, 1997);群落能量的现存量(standing crop of energy)体现一个群落在现有生态条件下所固定的太阳能总量,体现了群落在现有生态条件下的适应度,是群落生态学研究的重要内容,是对一群落进行评价的重要指标也是生态系统研究的重要内容。
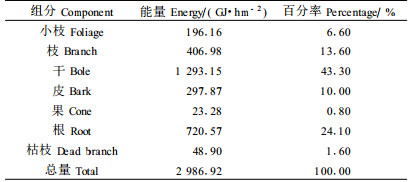
根据木麻黄试验样地生物量和各组分热值的测定结果,推算出样地各组分能量的现存量。从表 7可知:林分中能量现存量为2 986.92 GJ·hm-2,其中干最大,1 293.15 GJ·hm-2,占总量的43.30%,果最小23.28 GJ·hm-2,占总量的0.08%。具体各组分的大小顺序为:干(1 293.15 GJ·hm-2)>根(720.57 GJ·hm-2)>枝(406.98 GJ·hm-2)>皮(297.87 GJ·hm-2)>小枝(196.16 GJ·hm-2)>枯枝(48.90 GJ·hm-2)>果(23.28 GJ·hm-2)。整个林分的能量现存量比南亚热带鹤山大叶相思(Acacia auriculiformis)人工林的1 722 GJ·hm-2和马占相思人工林的2 782 GJ·hm-2(肖文发等,1999;彭少麟,2003)及南亚热带福建东山均一性风积沙土上的木麻黄人工林的1 057.51 GJ·hm- 2(叶功富等,2003)均高,但是比红壤性风积沙土木麻黄人工林的3 177.95 GJ·hm-2 (叶功富等,2003)低。可见在不同的土壤类型上,木麻黄能量现存量存在较大差异;在相同的土壤条件下,纬度和小气候不同,木麻黄能量现存量也存在较大差异(叶功富等,2003)。这主要是由于不同的土壤和小气候对木麻黄生长的养分和水分供养能力不同,造成木麻黄对光能的利用率的差异。
|
|
能量净固定量(net energy production,NEP)是指1年内单位面积上群落通过光合作用所固定的能量,扣除群落该时期因呼吸及被动物啃食等消耗的能量之后所剩余的能量(彭少麟等,1998)。其计算方法是以林分各组分(包括当年更迭的凋落物各组分)的年净干物质生产量和相应组分的干质量热值相乘之和而得。森林生态系统是一个开放的系统,与外界存在着强烈的物质和能量的交换,能量是任何系统的动力, 以前用生物量来衡量一个生态系统的生产力,存在着一定的不足,但从能量的角度可以更好地反映一个生态系统生产力水平。
通过对木麻黄生物量、生产力和各组分热值的测定,来计算木麻黄人工林生态系统生产力。木麻黄人工林能量年净固定量为196.80 GJ·hm-2a-1,干最大,109.82 GJ ·hm-2a-1, 占总量的55.81%, 果最小,2.43 GJ·hm-2a-1,占总量的1.23%。各组分能量净固定量大小顺序是:干(55.81%)>根(14.41%)>枝(10.36%)>小枝(9.96%)>皮(8.23)>果(1.23%)。
3.7 能量归还量生态系统从环境摄取能量的同时也向环境释放能量,在植物生态系统中,植物体主要通过凋落物向环境归还能量。
木麻黄林凋落物量和凋落物热值的乘积等于能量归还量。从表 8可知木麻黄凋落物能量年归还总量为294.56 GJ·hm-2a-1,其中小枝215.80 GJ·hm-2a-1,占总量的73.26%,枝条66.06 GJ·hm-2a-1,占22.43%,果12.92 GJ·hm-2a-1,占4.39%,花5.02 GJ·hm-2a-1,占1.07%。
|
|
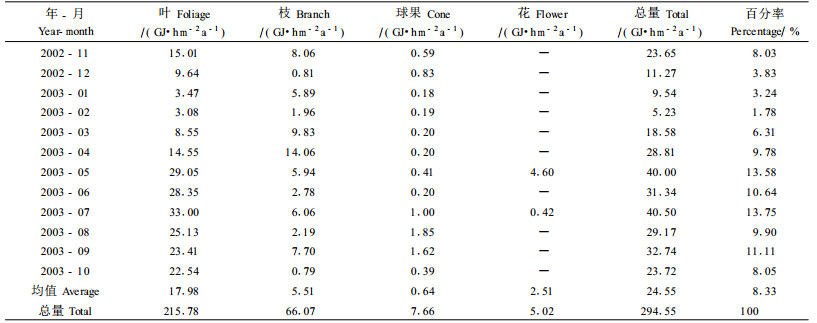
木麻黄林各组分各月份凋落物能量的归还量为294.56 GJ·hm-2a-1,叶归还量为2 15.80 GJ·hm-2a-1,占总量的72.99%,各月份的能量归还情况为:7月份最大,33.01 GJ·hm-2a-1,占叶总量的15.30%,2月份最小3.08 GJ·hm-2a-1,占总量的1.43%,各月份的情况为:7月份(15.30%)>5月份(13.46%)> 6月份(13.14%)>8月份(11.65%)>9月份(10.85%)>10月份(10.45%)>11月份(6.96%)>4月份(6.74%)>12月份(4.47%)>3月份(3.96%)>1月份(1.61%)>2月份(1.43%)。木麻黄小枝能量归还高峰期为6—8月,7月份最高,6—8月雨水充足,太阳辐射强,生长快,代谢也快。总凋落物归还情况变化与小枝相似,与凋落物中小枝占主体地位有关;枝条能量归还量在1年内变化不大,与小枝相比归还量小,这与枝条的生长周期长于小枝有关。
3.8 太阳能转化率群落的太阳能转化效率(energy conversing efficiency, ECE)是指群落每年净固定能量(NEP)占当年林地太阳辐射能(solar radiation energy, SRE)的百分比, 即ECE%=NEP/SRE×100。由于并非所有的太阳辐射能都可以被植物光合作用利用,一般用太阳光合有效辐射能(photosynthetic active radiation, PAR)代表群落接受到的总能量来计算群落的太阳能转化效率, 即ECE=NEP/PAR×100%。光合有效辐射能和太阳总辐射能的换算系数采用0.47,即PAR=SRE×0.47,按此公式计算(林鹏等,1999)。惠安崇武镇近20年的平均太阳辐射强度为4.62 GJ·hm-2a-1(数据来源于惠安气象站),根据木麻黄人工林能量的净固定量196.80 GJ·hm-2a-1,按上述公式计算,计算得木麻黄太阳能转化率为0.90%。能量净固定量要比武夷山黄山松的206.54 GJ·hm-2a-1 (林鹏等,1999)小,太阳能转化率比东北落叶松(Larix gmelinii)天然林的0.38%(肖文发等,1999)高。
4 小结1) 滨海沙地木麻黄人工林的现存生物量为152.60 t·hm-2,其中干占43.94%,根占24.14%,枝占13.74%,皮占9.40%,叶占6.42 %,枯枝占1.62%,果占0.76%。干和根有较高的生物量,占总量的68.08%,木麻黄林分高的树干和根系含量有利于提高抗台风的能力。
2) 滨海沙地木麻黄净生产力为10.17 t·hm-2a-1,其中干占总量的56.24%,根占总量的14.94%,两者共占总量的70.18%,说明木麻黄干和根具有较高的生产力,尤其是木麻黄发达的根系为木麻黄生长提供了良好条件。凋落物的归还量为14.17 t·hm -2a-1,占总生物量的9.27%,体现出木麻黄在沿海贫瘠的沙地上有很好的生长适应性和抵抗台风的能力。
3) 木麻黄各组分热值在19.26~20.53 kJ·g-1之间,整体平均值为19.70 kJ ·g-1,可知木麻黄有着较高的热值,具有较高的太阳能利用率,叶和皮热值较高,与前人的研究结果相似。木麻黄各组分灰分含量为1.20%~11.92%。
4) 林分能量现存量为2 986.92 GJ·hm-2,能量的净固定量为196.80 GJ·hm -2,能量的归还量为294.56 GJ·hm-2,有效太阳能转化率为0.90%。木麻黄人工林有着高的林分能量现存量、能量净固定量和太阳能的转化率,体现木麻黄具有高生产力,同时也反映出木麻黄在沿海退化沙地上具有很好的适应性。同时木麻黄有着较高的能量归还量,为木麻黄在滨海沙地上可持续生长提供了保障,可见木麻黄是东南沿海沙地很好的造林树种,现已成为沿海防护林的当家树种,具有不可替代的作用。
管东生. 1998. 香港桃金娘灌木群落植物生物量和净第一性生产量. 植物生态学报, 22(4): 356-363. DOI:10.3321/j.issn:1005-264X.1998.04.010 |
林鹏, 林益明, 李振基, 等. 1999. 武夷山黄山松群落能量的研究. 生态学报, 19(4): 504-507. DOI:10.3321/j.issn:1000-0933.1999.04.013 |
林鹏, 邵成. 1995. 福建亚热带雨林凋落物能流的研究. 厦门大学学报:自然科学版, 34(6): 987-991. |
林益明, 杨志伟, 李振基. 2001. 武夷山常绿林研究. 厦门:厦门大学出版社: 42-48; 58-63;124-127. |
柳江, 洪伟, 吴承祯, 等. 2002. 天然更新檫木林的生物量和生产力. 热带亚热带植物学报, 10(2): 105-110. DOI:10.3969/j.issn.1005-3395.2002.02.002 |
刘玉萃, 吴明作, 郭宗民, 等. 1998. 宝天曼自然保护区栓皮栎林生物量和净生产力研究. 应用生态学报, 9(6): 569-574. DOI:10.3321/j.issn:1001-9332.1998.06.002 |
彭少麟. 2003. 热带亚热带恢复生态系统研究与实践. 北京: 科学出版社.
|
彭少麟, 任海. 1998. 南亚热带森林生态系统的能量生态研究. 北京: 气象出版社, 45-60, 74-79.
|
孙雪峰, 陈灵芝. 1997. 暖温带落叶阔叶林能量分配的综合特征, 暖温带森林生态系统结构与功能研究. 北京: 科学出版社, 163-172.
|
温达志, 魏平, 孔国辉, 等. 1997. 鼎湖山锥栗+黄果厚壳桂+荷木群落生物量及其特征. 生态学报, 17(5): 497-504. DOI:10.3321/j.issn:1000-0933.1997.05.008 |
吴寿德, 叶功富, 潘惠忠, 等. 1996. 木麻黄人工林生物产量的结构特征. 防护林科技, (专刊): 21-24. |
肖文发, 徐德应. 1999. 森林能量利用与产量形成的生理生态基础. 北京: 中国林业出版社.
|
叶功富, 林银森, 吴寿德, 等. 1996a. 木麻黄林生产力动态变化的研究. 防护林科技, (专刊): 17-20. |
叶功富, 隆学武, 潘惠忠, 等. 1996b. 木麻黄林的凋落物动态及其分解. 防护林科技, (专刊): 30-34. |
叶功富, 张清海, 林益明, 等. 2003. 海岸带不同立地木麻黄群落能量研究. 林业科学, 39(专刊): 1-7. |
蚁伟民, 张祝平, 丁明懋, 等. 2000. 鼎湖山格木群落的生物量和光能利用效率. 生态学报, 20(2): 397-403. |
祖元刚. 1990. 能量生态学. 长春: 吉林科技出版社.
|
Ares A, James H F. 2000. Productivity, nutrient and water-use efficiency of Eucalyptus asligna and Toona ciliata in Hawaii. Forest Ecology and Management, 139: 227-236. |
Gupta S K. 1972. Energy structure of standing crop in certain grasslands at Gyanpur. Trop Ecol, 13: 147-155. |
Jordan C F. 1971. Productivity of a tropical forest and its relation to a world pattern of energy storage. J Ecol, 59: 127-142. DOI:10.2307/2258457 |
McConnaughay K D M, Coleman J S. 1999. Biomass allocation in plants. Ecology, 80(8): 2581-2593. |
Labrecque M, Teddorescu T I, Dajgle S. 1997. Biomass productivity and wood energy of Salix species after 2 years growth in sric fertilized with wastewater sludge. Biomass and Bioenergy, 12(6): 409-417. DOI:10.1016/S0961-9534(97)00011-1 |
Lodhiyal N, Lodhiyal L S. 2003. Biomass and net primary productivity of Bhabar shisham forests in cental Himalaya, India. Forest Ecology and Management, 176: 217-235. DOI:10.1016/S0378-1127(02)00267-0 |
Odum H T. 2002. Explanations of ecological relationships with energy systems concepts. Ecological Modelling, 158: 201-211. DOI:10.1016/S0304-3800(02)00232-6 |
Strivastava A K. 1995. Biomass and energy production in Casuarina equisetifolia plantation stands in the degraded dry tropic of the Vindhyan Platean, India. Biomass and Bioenergy, 9(6): 465-471. DOI:10.1016/0961-9534(95)00048-8 |