 2006, Vol. 42
2006, Vol. 42文章信息
- 李新岗, 刘惠霞, 刘拉平, 马养民.
- Li Xingang, Liu Huixia, Liu Laping, Ma Yangmin.
- 影响松果梢斑螟寄主选择的植物挥发物成分研究
- Study on Host-Plant Volatiles Affecting the Host Selection of Dioryctria pryeri
- 林业科学, 2006, 42(6): 71-78.
- Scientia Silvae Sinicae, 2006, 42(6): 71-78.
-
文章历史
- 收稿日期:2005-01-20
-
作者相关文章


2. 西北农林科技大学植物保护学院 杨凌 712100;
3. 西北农林科技大学测试中心 杨凌 712100
2. College of Plant Protection, Northwest Agriculture Forestry Science and Technology University Yangling 712100;
3. Testing Centre, Northwest Agriculture Forestry Science and Technology University Yangling 712100
近年来, 植物挥发物对植食性昆虫寄主选择影响的研究越来越多(娄永根等, 2000), 特别是对鳞翅目昆虫的研究发现:单食性或寡食性蛾类主要依靠昆虫嗅觉和植物挥发物进行寄主选择(Ramaswamy, 1988)。根据成虫取食先于产卵的特性, 针叶树球果害虫产卵选择主要有3种假设模式:一是成虫取食球果的害虫, 寄主树信号(挥发物)在中远距离定向上起作用, 而球果信号则在近距离适合度或维持上起作用(Mattson et al., 1986; Turgeon et al., 1994); 二是成虫取食花或花蜜的球果害虫, 其长距离的寄主定向主要利用寄主树和球果信号, 一旦成虫到达树冠上, 其球果定位和球果适合度主要依靠球果信号(Turgeon et al., 1994); 三是成虫对中、长距离的球果识别主要依靠视觉和嗅觉的结合(Rappaport et al., 1998; Turgeon et al., 1994)。关于梢斑螟属(Dioryctria)球果害虫的研究认为, 雌成虫主要依靠挥发性萜类成分进行寄主和球果定向(Fatzinger et al., 1985; Hanula et al., 1985;Shu et al., 1997), 但具体影响球果螟成虫寄主选择和越冬幼虫转移危害的机制还不清楚。
梢斑螟属是针叶树球果害虫的一大类, 种类多, 竞争性强, 危害严重(Yates Ⅲ, 1986; Spies, 1985; 李新岗, 2002)。松果梢斑螟(Dioryctria pryeri)是我国松类球果和枝梢的主要害虫, 属寡食性害虫, 在我国北方严重危害油松球果(李宽胜, 1992)。松果梢斑螟1年1代, 其寄主选择有2个关键环节, 一是7、8月份成虫产卵选择, 二是翌年5月份越冬幼虫转移危害和选择。为了探明松果梢斑螟寄主选择的机理, 本文针对其转移危害和成虫产卵的寄主选择, 研究其寄主挥发物成分的差异及其对松果梢斑螟寄主选择的影响, 为该虫综合管理提供理论基础。
1 材料与方法 1.1 材料来源油松(Pinus tabulaeformis)、华山松(P. armandi)健康球果、虫害果以及针叶均采自陕西陇县八渡油松良种基地。油松种子园营建于1975年, 海拔1 100~1 300 m, 树高约6 m, 375株·hm-2, 郁闭度0.5~0.6, 坡度20°~30°。华山松树龄25~30 a, 树高约5 m, 郁闭度0.5~0.6, 坡度20°~30°, 在油松种子园外围。
球果、针叶采样时间分别在松果梢斑螟幼虫转移危害期的5月上中旬和成虫产卵期的7月份。由于样品离体后挥发物成分容易发生变化, 因此, 采样时需带枝采样, 并尽快对样品进行浸提或吸附处理(Turgeon et al., 1998)或进行幼虫选择试验。室内趋向性试验选用2~3龄饥饿2 h的松果梢斑螟幼虫, 于2004年5月中旬采自陇县油松种子园的虫害球果, 带回实验室待用。
1.2 球果及浸提液趋向性测试 1.2.1 幼虫对不同类别球果的趋向性反应在直径15 cm, 高5 cm的培养皿中, 分别在3个方向放入健康的油松球果和油松球果小卷蛾(Gravitarmata margarotana)危害果、油松虫害果和华山松虫害油松球果(小卷蛾危害)果, 以及华山松虫害果和健康果; 每组进行20~30个重复, 分别在5、10、30、60 min, 1和3 h记录球果螟幼虫在各类球果上的位置, 每出现1次记录1次, 在空白处记0, 统计各类球果上的出现频率, 反映球果螟的趋向反应; 并同时记录其蛀果情况。由于松果梢斑螟幼虫在10—16时为出果活动期, 所以试验一般在幼虫钻蛀球果的17—19时进行。
1.2.2 幼虫对球果提取液的趋向性反应在直径15 cm、高5 cm的培养皿中, 先放上一片与培养皿等大的白纸; 用直径7 cm的半块滤纸, 分别滴上:1)油松健康果、油松球果小卷蛾危害果二氯甲烷浸提液, 以及二氯甲烷液为对照; 2)松果梢斑螟危害果、油松球果小卷蛾危害果和健康果(对照)二氯甲烷浸提液, 共2组6种处理的滤纸, 待二氯甲烷挥发、滤纸风干后, 在培养皿内3个方向, 随即放上处理滤纸, 每次试虫1头, 放于培养皿正中央, 盖上盖子, 5 min后观察幼虫的选择。每个培养皿试虫1头, 5个1组, 重复4~5次(Hughes et al., 2003)。数据进行平方根处理, 然后比较差异显著性。
1.3 植物挥发物分析根据Dormont等(1997)的研究, 针叶树球果、针叶浸提法与XAD2吸附法获得的萜类挥发物基本一致。本实验采用了二氯甲烷溶剂提取和XAD2吸附2种方法。
1.3.1 样品浸提幼虫转移期, 选取油松2个家系各2株样树; 成虫产卵期, 只选取1个家系(F1)2株样树。2个时期, 华山松均选取1个家系2株样树。因为松类结果, 树冠中部最有代表性(李宽胜, 1992), 阳面由于温度高而球果或针叶的挥发强度大, 阴面的挥发强度较小(李金龙等, 1994)。所以, 采样选取树冠中部半阳面结果枝。每个处理在2株样树东面和西面共选取4个样品, 任选3个为重复。幼虫期每个样品有油松球果4~5个, 华山松球果3个, 其中健康球果约50 g, 虫害果约30 g, 分别用二氯甲烷(色谱纯, 美国TEDIA公司)50 mL和30 mL; 成虫产卵期每个样品有球果3~4个, 其中健康果约50 g, 虫害果约30 g, 当年生针叶20 g, 分别用二氯甲烷50、30和20 mL, 浸提24 h, 过滤, 置于-20 ℃以下冰箱中待用(Dormont et al., 1997)。
1.3.2 球果挥发物吸附选择油松2个家系各2株样树, 每个样树在树冠中部东西两面, 分别选取健康球果和小卷蛾虫害球果各4个样品, 任选3个为重复; 每个样品枝有球果2个(同为虫害果或健康果), 带枝活体采样; 吸附剂选用XAD2, 活化后烘干装柱, 每柱装吸附剂1.5 g, 吸附30 min, 空气流速1 L·min-1; 随后用3 mL二氯甲烷洗脱2 h, 样品置于-10 ℃以下冰箱中待分析(Dormont et al., 1997)。
1.3.3 GC-MS分析挥发物的鉴定利用美国Thermo-Finnigan公司的TraceDSQ气相色谱-质谱仪(GC-MS)(西北农林科技大学测试中心)分析, 色谱柱利用SE-54毛细管柱(30 m×0.25 mmID, 膜厚0.25 μm), 载气氦气; 流速1 mL·min-1, 进样1 μL, 分流比50:1。起始温度40 ℃, 停留2.5 min, 以8 ℃·min-1升到250 ℃。质谱条件:EI离子源, 电离能70 eV。各成分通过与谱库(NIST2002版)标准化合物的质谱图核对并分析后, 进行定性, 根据峰面积归一化法进行定量。
1.4 统计分析统计分析采用ExcelStat (Installshield Software Corporation)分析软件。采用单因素(ANOVA) F检验(Fisher LSD post-hoc tests)比较不同处理间的差异显著性。P<0.05, 不同处理间的差异显著性用a、b、c标注, 不标注表示无显著差异。图、表中的数据为平均值±SE, n代表样本重复数。
2 结果与分析 2.1 室内活性测定 2.1.1 幼虫对不同类别球果的趋向性反应图 1表明:在油松健康果和虫害果选择中, 梢斑螟幼虫明显趋向虫害果, 出现频率96.7%, 并全部蛀食虫害果; 而在油松虫害果与华山松虫害果的选择中, 幼虫明显趋向油松虫害果, 出现频率高达97%, 蛀果也全部是油松虫害果; 在华山松虫害果和健康果的选择中, 幼虫的趋向性不明显, 即使在2类球果上, 也不蛀食华山松球果。

|
图 1 松果梢斑螟幼虫对健康和虫害的油松和华山松球果的趋向和选择反应 Fig. 1 Trend and selection responses of pine conewormlarvae to uninfested and infested cone of P. tabulaeformis and P. armandi A1/A2油松虫害果/健康果Infested/uninfested cone of P. tabulaeformis (Pt) (n=30); B1/B2油松虫果/华山松虫果Infested cone of Pt/ P. armandi (Pa)(n=30); C1/C2华山松虫果/健康果Infested/uninfested cones of Pa(n=20). |
图 2表明:在油松健康球果和小卷蛾危害果浸提液的趋向反应中, 趋向小卷蛾危害果浸提液的幼虫数量(4.4头)显著高于趋向健康果浸提液的幼虫数(0.6)和二氯甲烷对照(0)。在油松健康果、小卷蛾危害果和梢斑螟危害果的选择中, 也是显著趋向小卷蛾危害果(3.2头), 趋向梢斑螟危害果提取液的幼虫只有1.4头, 趋向健康果浸提液对照为0.4头。总之, 松果梢斑螟幼虫明显趋向虫害的油松球果浸提液。

|
图 2 松果梢斑螟幼虫对不同油松球果提取物的趋向性反应(2004年5月) Fig. 2 Trend responses of pine coneworm larvae to extract solvent of uninfestedand infested cone by G. margarotana and D.pryeri (May, 2004) CK:二氯甲烷溶剂CH2Cl2 solvent; H:健康球果提取液Extract solvent of uninfested cone with CH2Cl2; G:小卷蛾危害果提取液Extract solvent of infested cone by G. margarotana larvae with CH2Cl2; D:球果梢斑螟危害果提取液Extract solvent of infested cone by D. pryeri larvae with CH2Cl2. *代表处理间的差异显著性Asterisks indicate level of significant differences among treatments and control. * P<0.05; ** P<0.01。Treatment CK, H, G, n=5; Treatment G, D, H, n=4 |
表 1结果表明, 吸附法得到油松球果单萜化合物7种, 2个家系球果挥发物成分基本一致, 但各成分的含量差异较大, 其中6种成分含量差异显著。2号家系球果挥发成分含量最多的是β-香叶烯, 含量为73.7%, 其次是α-蒎烯, 为13.72%, β-蒎烯的含量也显著高于1号家系。1号家系球果虫害后, 4种组分含量显著变化, 其中α-蒎烯和β-香叶烯含量显著减少, β-非兰烯和β-蒎烯含量显著增加, 而D-柠檬烯成分没有明显变化, 莰烯和对-薄荷-1(7), 3-二烯也均处于痕迹含量水平, 没有变化。
|
|
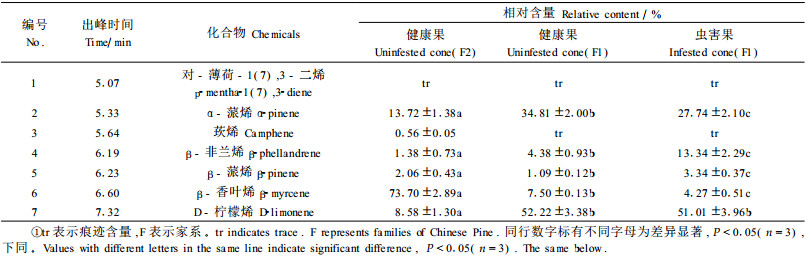
油松球果二氯甲烷浸提液的GC-MS分析结果如表 2所示:共检出挥发性单萜成分12种, 比吸附法检出的多5种, 分别是3-侧柏烯、δ-萜品油烯、异松油烯、乙酸龙脑酯和乙酸天竺葵酯, 其中前2种成分为痕迹含量, 后3种成分含量较多, 但出峰时间较迟。差异显著性比较发现, 虫害后特异性的单萜组分为α-蒎烯、β-非兰烯、β-香叶烯、D-柠檬烯和异松油烯, 以及乙酸龙脑酯和乙酸天竺葵酯。其中, 前4种组分与吸附法的结果一致。由于溶剂浸提可将球果组织松脂道中的贮存萜类提取出来(Dormont et al., 1997; Pureswaran et al., 2004), 因而浸提结果在组成和含量上与吸附结果有一定差异, 但主要挥发成分基本一致。虫害后, 1号家系球果单萜显著增加的成分是α-蒎烯、D-柠檬烯和乙酸龙脑酯, 显著减少的组分是β-香叶烯; 虫害后, 2号家系显著增加的成分是α-蒎烯和乙酸龙脑酯, 其他显著变化的组分均明显减少。
|
|
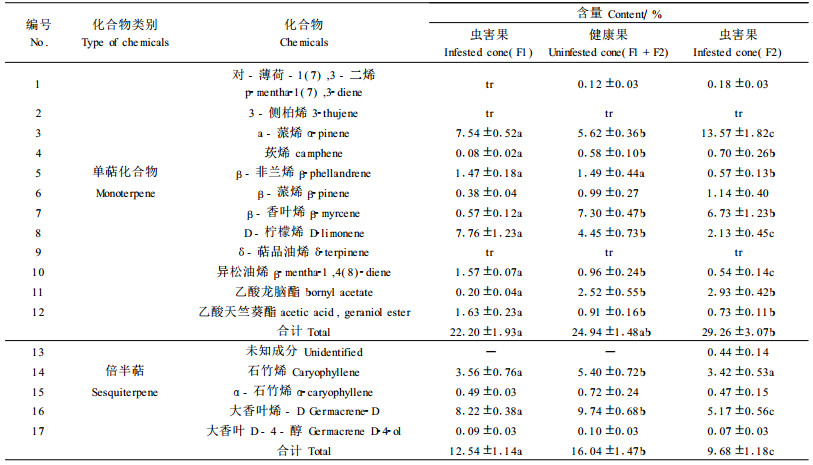
表 2也可看出:油松球果受害后, 2个油松家系各倍半萜成分和提取液中倍半萜的含量都有所下降, 但主要成分都是大香叶烯-D、石竹烯和α-石竹烯。在峰16和17之间, 还有3种痕迹含量成分, 分别是α-木罗烯(α-muurolene)、1, 5二甲基-8-异丙基-1, 5-环十(烷)二烯[1, 5-dimethyl-8-(1-methylethyl)-1, 5-cyclodecadiene]和杜松-1(10), 4-二烯[cadin-1(10), 4-diene], 其含量都低于0.08%, 并且在3类球果中的含量变化不大。
2.2.3 华山松球果二氯甲烷提取液中的萜类成分表 3为华山松球果提取液中的萜类挥发物成分。华山松球果浸提液中共检出单萜成分12种(11个峰位), 其中β-非兰烯和4-侧柏烯出现在同一个峰上, 不能分开; 在健康球果中, 有5种痕迹含量成分; 含量较多的是D-柠檬烯、β-香叶烯、β-蒎烯、β-非兰烯/4-侧柏烯、α-蒎烯和异松油烯。与油松相比, 华山松球果提取液中单萜含量较高, 并且球果虫害后, 除D-柠檬烯略有增加外, 各主要单萜成分和总含量都有所下降, 其中β-蒎烯、β-香叶烯和单萜总含量显著减少; 而微量成分β-罗勒烯、r-萜品烯和乙酸龙脑酯含量有所增加; 3-侧柏烯和莰烯一直保持痕迹含量, 没有变化。
|
|

与油松球果浸提液中单萜成分比较, 化合物数量都是12种, 但有5种化合物成分不一致。华山松球果少了对-薄荷-1(7), 3-二烯和乙酸天竺葵酯, 增加了β-罗勒烯和4-侧柏烯(在β-非兰烯峰位), 油松球果中的δ-萜品油烯在华山松球果中变成r-萜品烯。华山松健康球果中, 5个主峰位6个主要成分(a-蒎烯、β-非兰烯/4-侧柏烯、β-蒎烯、β-香叶烯和D-柠檬烯)的含量均高出5%, 最多的是D-柠檬烯, 16.92%;而油松健康球果除了家系差异外, 超过或接近5%的化合物只有3个, 分别是α-蒎烯、β-香叶烯和D-柠檬烯。
表 3也可看出:华山松球果虫害后, 各倍半萜成分和提取液中倍半萜的含量都有所增加, 其中新增加了4种含量较少的倍半萜成分; 主要成分只有石竹烯1种。与油松相比, 不论是健康球果还是虫害果, 倍半萜成分差异较大。
2.3 成虫产卵期油松球果、针叶中萜类挥发物成分 2.3.1 单萜成分表 4列出了松果梢斑螟雌成虫产卵选择期油松各部位提取液中的单萜成分。健康球果二氯甲烷浸提液共检出挥发性单萜成分13种, 除β-非兰烯峰位有4-侧柏烯外, 与幼虫转移期健康球果单萜成分基本一致; 在含量上, 则略有差异。α-蒎烯含量最多(11.29%), 其次是β-香叶烯和D-柠檬烯(分别是9.13%和3.94%); β-蒎烯的含量只有0.73%, 乙酸龙脑酯和乙酸天竺葵酯的含量与幼虫转移危害期不同, 分别是0.93%和0.30%, 原来的δ-萜品油烯变成r-萜品烯, 含量0.21%。
|
|
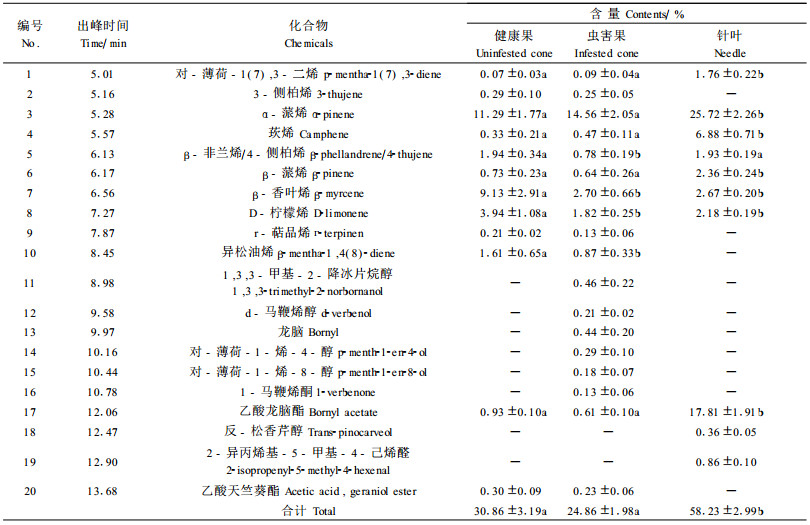
此时的虫害果萎缩干枯, 呈红褐色, 检出挥发性单萜峰位17个, 化合物18种。球果受害后, α-蒎烯含量有所增加, 而β-非兰烯/4-侧柏烯、β-蒎烯、D-柠檬烯和异松油烯含量显著减少; 并新增加6种成分, 含量都比较低, 其中d-马鞭烯醇和1-马鞭烯酮是α-蒎烯的衍生物, 可能与害虫排泄物有关(Pureswaran et al., 2004); 单萜总含量有所减少。
针叶中检出峰位11个(分2个峰区出现), 单萜化合物12种。主峰区各种单萜成分含量较高, 均超过1.5%, 其中含量最高的是α-蒎烯(25.72%), 其他成分的含量与健康果和虫害果差异较大。莰烯含量达到6.88%, 而β-香叶烯和D-柠檬烯含量较低, 分别为2.67%和2.18%。次峰区共3种成分, 其中乙酸龙脑酯的含量最多, 为17.81%, 显著高于2类球果。
2.3.2 倍半萜成分如表 5所示, 健康球果、虫害果和针叶二氯甲烷提取液中, 倍半萜含量分别为46.18%、24.76%和23.1%, 健康球果的含量显著高于虫害果和针叶。健康球果和虫害果中倍半萜成分一致, 均为14种; 3种主要成分也一致, 分别是未知成分、石竹烯和α-石竹烯。球果受害后, 6种成分显著变化, 其中, 未知成分、石竹烯、α-石竹烯和古巴烯含量显著减少, 而十氢-4, 8, 8-三甲基-9-亚甲基-1, 4-亚甲基奥和衣兰烯含量显著增加。
|
|

与健康球果和虫害果相比, 针叶提取液中倍半萜成分变化较大(表 5), 共有11种成分; 含量最多是石竹烯, 其含量显著高于2类球果; 未知成分的含量也显著不同于2类球果; α-木罗烯和桉叶烷-4(14), 11-二烯为针叶中的特有成分, 含量较低, 而球果中的α-长叶松烯、衣兰烯、杜松-1(10), 4-二烯等5种成分在针叶中没有检出。
3 结论与讨论 3.1 结论松果梢斑螟幼虫明显趋向和选择油松球果而不选择华山松球果; 在油松球果及其提取液试验中, 幼虫则明显趋向和选择虫害球果; 成虫也明显趋向和选择虫害油松球果产卵(待发表)。这表明小卷蛾虫害果对松果梢斑螟幼虫和成虫均有引诱作用。进一步研究表明:梢斑螟幼虫转移期和成虫产卵期, 油松健康球果、虫害果及针叶挥发性萜类成分显著不同。虽然同一时期的虫害果和健康果挥发性主要组分一致, 但球果受害后, 各组分含量明显变化。幼虫转移期球果虫害后, α-蒎烯、β-蒎烯、β-非兰烯、β-香叶烯、D-柠檬烯、异松油烯、乙酸天竺葵酯、大香叶烯-D和石竹烯等为特异性成分; 而成虫产卵期的虫害球果, α-蒎烯、β-非兰烯/4-侧柏烯、β-香叶烯、D-柠檬烯、未知成分、石竹烯和α-石竹烯等为特异性萜类成分(表 4, 表 5), 它们可能对松果梢斑螟具有引诱作用, 其具体作用有待进一步的研究。2个时期华山松健康果和虫害果挥发物组成和含量与油松球果显著不同(李新岗等, 2005), 并且单萜挥发物含量多, 倍半萜含量少。
3.2 讨论在林间条件下, 松果梢斑螟也可以危害华山松(李宽胜, 1992), 但在本试验的油松种子园条件下, 该虫明显趋向和选择油松, 这表明油松为最适宜寄主。并且, 松果梢斑螟雌成虫产卵主要在虫害的干枯或半干枯油松球果及其附近的当年生针叶叶鞘处, 几乎不选择健康的球果(李宽胜, 1992), 这与美洲冷杉梢斑螟(D. abietivorella)和南方松球果螟(D. amatella)雌成虫的产卵选择情况基本一致(Fatzinger et al., 1985; Hanula et al., 1985; Fidgen et al., 1996)。南方松球果螟雌蛾明显趋向受害干枯、单萜挥发成分含量高的组织产卵, 不趋向没有虫害或单萜挥发成分含量低的寄主组织产卵, 其结论与松果梢斑螟的选择一致。进一步研究发现:α-蒎烯在南方松球果螟雌蛾产卵位置识别中起着重要的作用, 并且产卵很大程度上受到其他单萜成分的强化, 如香叶烯和柠檬烯(Fatzinger et al., 1985; Hanula et al., 1985)。相比之下, 梢斑螟成虫产卵期, 油松球果虫害后挥发物强度增加, 特别是α-蒎烯、β-非兰烯/4-侧柏烯、β-香叶烯、D-柠檬烯和石竹烯等可能与成虫的寄主选择有关(表 4), 而华山松球果上的特异成分可能具有驱避作用, 但其具体作用还需昆虫触角电位(EAG)测试和生测结果的证实。
虫害诱导的植物挥发物引诱昆虫天敌, 驱避同种或异种昆虫的产卵; 也有虫害诱导的植物挥发物引诱同种或异种昆虫的产卵(娄永根等, 2000)。松果梢斑螟趋向小卷蛾虫害果, 而虫害后油松球果抗营养、抗消化酶类增加(直接防御), 天敌被引诱(间接防御)(待发表), 表明虫害诱导的防御已被梢斑螟克服。梢斑螟属昆虫多属此类, 所以作者推测:寄主松树虫害组织单萜挥发物可能在成虫的远距离寄主定向中起作用(Fatzinger et al., 1985; Hanula et al., 1985)。而本研究的结论可以推断:低挥发性的倍半萜以及果实特性可能在近距离的适合度上起作用; 虫害球果挥发物对幼虫的吸引主要在近距离范围内起作用, 而球果营养可能在球果适合度上起作用(Chapman, 2003)。幼虫转移危害期和成虫产卵期球果的发育阶段不同, 球果挥发性萜类成分不同, 其虫害诱导后的挥发物也不同, 因此推测, 吸引成虫和幼虫的挥发成分组成是不同的, 但主成分可能是一致的(Dickens, 2002)。
李宽胜. 1992. 油松种实害虫防治技术研究. 西安: 陕西科学技术出版社.
|
李新岗, 等. 2002. 油松球果害虫的危害机理研究. 西北农林科技大学学报, 30(2): 77-82. |
李新岗, 马养民, 刘拉平, 等. 2005. 华山松球果挥发性萜类成分研究. 西北植物学报, 25(10): 2072-2076. DOI:10.3321/j.issn:1000-4025.2005.10.024 |
李金龙, 白郁华, 胡建信, 等. 1994. 油松排放萜类化合物浓度的日变化及排放速率的研究. 中国环境科学, 14(3): 165-169. |
娄永根, 程家安. 2000. 虫害诱导的植物挥发物:基本特征、生态学功能及释放机制. 生态学报, 20(6): 1097-1106. DOI:10.3321/j.issn:1000-0933.2000.06.031 |
Chapman R F. 2003. Contact chemoreception in feeding by phytophagous insects. Annu Rev Entomol, 48: 455-484. DOI:10.1146/annurev.ento.48.091801.112629 |
Dickens J C. 2002. Behavioural responses of larvae of Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae), to host plant volatile blends attractive to adults. Agricultural and Forest Entomology, 4(4): 309-314. DOI:10.1046/j.1461-9563.2002.00153.x |
Dormont L, Roques A, Malosse C. 1997. Efficiency of spraying mountain pine cones with leoresin of swiss stone pine cones to prevent insect attack. J Chem Ecol, 23(10): 2261-2274. DOI:10.1023/B:JOEC.0000006672.05861.6c |
Fatzinger C W, Merkel E P. 1985. Oviposition and feeding preferences of the southern pine coneworm for different host-plant-materials and observations on monoterpenes as an oviposition stimulant. J Chem Ecol, 11(6): 689-699. DOI:10.1007/BF00988298 |
Fidgen L, Sweeney J. 1996. Fir coneworms, Diorytria abietivorella, prefer cons previously exploited by spruce cone maggots, Strobilomyia neatithracina Michelsen and S. appalachensis Michelsen. IUFRO Working Party: S7.03-01Cone and Seed Insects, 5-6
|
Hanula J L, Berisford C W, Debarr G L. 1985. Monoterpene oviposition stimulants of Diorytria amatella in volatiles from fusiform rust galls and second-year loblolly pine cones. J Chem Ecol, 11(7): 943-952. DOI:10.1007/BF01012080 |
Hughes W O, Galley D, Knapp J K. 2003. Host location by adult and larval codling moth and the potential for its disruption by the application of kairomones. Entomol Exp Appl, 106: 147-153. DOI:10.1046/j.1570-7458.2003.00020.x |
Mattson W J, Strauss S H. 1986. Are host volatiles involved in cone finding by Conophthotus resinosae? Proc Cone and Seed Insect Working Party Conf (IUFROS2.07-01), 2rd, Brancon, 1986. Olivet: INRA, 185-204
|
Pureswaran D S, Regine G, Borden J H. 2004. Quantitative variation in monoterpenes in four species of conifers. Biochem System Ecol, 32: 1109-1136. DOI:10.1016/j.bse.2004.04.006 |
Ramaswamy S B. 1988. Host finding by moth: sensory modalities and behaviours. J Insect Physiol, 34: 235-249. DOI:10.1016/0022-1910(88)90054-6 |
Rappaport N G, Jenkins M J, Roques A. 1998. Cone and foliage volatiles from Douglas-fir and European larch: relationships to attack by cone ane seed insects//DeBarr G L, Roques A, Sun J H, et al. Proceedings of the 4th Cone and Seed Insects Working Party Conference (IUFROS2.07-01), Beijing and Harbin, 1992. Georgia: USDA Forest Service Southeastern Forest Experiment Station
|
Shu S, Grant G G, Langevin D A, et al. 1997. Oviposition and ecectroantennogram responses of Dioryctria abietivorella (Lepidoptera:Pyralidae) elicited by monoterpenes and enantiomers from estern white pine. J Chem Ecol, 23(1): 35-50. |
Spies C J. 1985. Some populational relationships between spruce budworm Choristoneura fumiferana and spruce coneworm Diorycteria reniculelloides. Can Entomol, 117: 193-202. DOI:10.4039/Ent117193-2 |
Turgeon J J, Brocherhoff E G. 1998. Difference in composition and release rate of volatiles emitted by black spruce seed and cones sampled in situ versus ex situ. Can J Forest Res, 28: 311-316. DOI:10.1139/x97-216 |
Turgeon J J, Roques A, Groot P. 1994. Insect fauna of coniferous seed cones: diversity, host plant interactions, and management. Annual Rev Entomol, 39: 179-212. DOI:10.1146/annurev.en.39.010194.001143 |
Yates Ⅲ H O. 1986. Checklist of insect and mite species attacking cones and seeds of world conifers. J Entomol Sci, 21(2): 142-168. DOI:10.18474/0749-8004-21.2.142 |