 2006, Vol. 42
2006, Vol. 42文章信息
- 余仲东, 刘莉丽, 曹支敏, 张星耀.
- Yu Zhongdong, Liu Lili, Cao Zhimin, Zhang Xingyao.
- 杨树溃疡病菌异核体现象的研究
- Study on Heterokaryotic Phenomenon of Poplar Canker Pathogen
- 林业科学, 2006, 42(5): 142-144.
- Scientia Silvae Sinicae, 2006, 42(5): 142-144.
-
文章历史
- 收稿日期:2004-09-29
-
作者相关文章


2. 中国林业科学研究院 北京 100091
2. Chinese Academy of Forestry Beijing 100091
异核体现象是真菌遗传变异和基因流动的基础,在小麦条锈菌毒性变异中起着重要的作用(康振生等, 1993)。真菌中广泛存在的营养体亲和现象,使异核体现象在真菌的进化、适应性、致病性变异中更具重大意义(余仲东等, 2002)。本文研究杨树溃疡病菌异核体现象的细胞学及分子生物学证据,及异核体现象在病菌致病性变异中的作用。
1 材料与方法 1.1 供试材料菌株A,从陕西杨凌的北京杨(Populus beijingensis)上分离获得。菌株B来自辽宁铁岭,从箭杆杨[P. nigro var. thevestina(Dode)]上分离获得。
1.2 异核体菌株的培养及培养特性用打孔器(Φ 0.5 cm)取PDA上培养5 d的A、B菌落,在基本培养基上对峙培养,待形成稳定的生长配接线后,取配接线菌丝体单独培养,获得单一菌落后,取菌丝顶端培养,直到获得稳定的异核体菌落,并记录菌株生长时的菌落特征。
1.3 细胞核染色采用Fulgen染色法对亲本、异核体菌株菌丝体细胞核染色,染色方法参照郑若立(1978)。
1.4 致病性测定将A、B菌株、异核体株系分别接种在红富士苹果上(Φ 9 cm),每苹果接2孔(孔深0.5 cm),每孔1个菌饼。设1个对照孔,不接菌饼。保湿培养,逐日记录垂直2方向病斑大小,取均值并减去对照孔直径作为病斑发病大小。每接种重复2次。将所得数据用软件SPSS(Version11.5,American)进行单因素方差分析和多重比较。
1.5 ITS-rDNA-RFLP分析按Zhu等(1993)方法提取DNA。采用通用引物ITS1、ITS4对核糖体基因转录间隔区进行扩增。引物序列为:ITS1 5′ > TCCGTAGGTGAACCTGCG < 3′、ITS4 5′ > TCCTCCGCTTATTGATAT < 3′。Taq多聚酶为鼎国生物工程公司生产。
反应体系50 μL。包括ddH2O 27 μL,10× Buffer 5 μL,Mg2+(25 μmol·L-1)3 μL,4dNTPs(10 mmol·L-1) 1 μL,ITS1(10 μmol·L-1) 2.5 μL,ITS4(10 μmol·L-1) 2.5 μL, Taqpolymerase(1 U·μL-1) 4 μL, DNA 5 μL,共计50 μL。反应条件:94 ℃预变性2 min,55 ℃停留40 s,93 ℃变性1 min,51 ℃退火30 s,72 ℃延伸1 min 30 s,35个循环,4 ℃终止反应。PCR产物用HhaⅠ(识别序列:GCG/C)和HpaⅡ(识别序列C/CGG)酶解,反应条件按说明书。酶解产物采用8%PAGE电泳,成像系统成像并分析结果。
2 结果与分析 2.1 异核菌株及A、B菌株培养生物学特性比较将A、B菌株对峙培养后,2菌落边缘菌丝相互交织,形成隆起的、气生菌丝发达的白色生长配接线,配接线的基部有显著的色素沉积。取配接线菌丝培养获得3株系(AB1、AB2、AB3)。菌落(AB)均具有菌落A和B的培养特征,3株系菌丝尖端培养形成的菌落,其培养特征近同(图 1)。

|
图 1 异核体菌株及亲本菌株培养生物学特性 Fig. 1 The culture characteristics of heterocarytic strains and their parental strains |
Fulgen染色结果表明:A菌株为单核菌株;B菌株为单核菌株或双核,以单核常见;AB为异核体菌株,具有双核或3核,以双核为主(图 2)。

|
图 2 菌丝体细胞核染色结果 Fig. 2 Dying results of nucleolus |
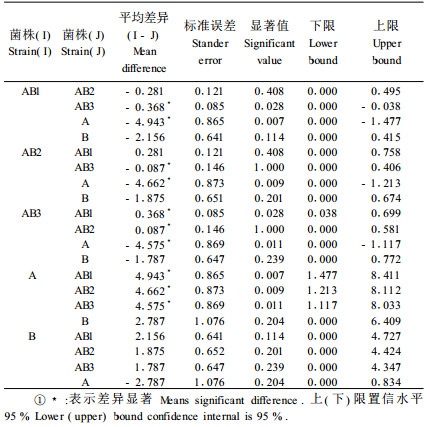
经单因素方差分析表明,供试菌株致病力大小存在显著差异(表 1)。异核体菌株平均致病力均低于亲本菌株致病力大小,其中AB1最小(图 3)。差异一致性检验表明,各菌株差异显著性不一致(表 2)。故采用Tamhane法进行多重比较。结果表明:异核体菌株平均致病力大小与亲本A菌株平均致病力差异显著,与B菌株平均致病力差异不显著。异核体菌株间,AB1与AB3平均致病力差异显著、与AB2平均致病力差异不显著、AB2与AB3平均致病力差异显著(表 3)。
|
|


|
图 3 异核体与亲本菌株平均致病力大小 Fig. 3 Mean patholognicity of heterokaryotic strains and parental |
|
|

|
|
PCR扩增ITS区段获得特异性600bp片段。经2种4碱基高频内切酶酶解,异核体菌株AB同亲本菌株A、B酶解格局无明显差异(图 4)。

|
图 4 ITS-rDNA-PCR及RFLP分析 Fig. 4 Products of ITS-rDNA-PCR and RFLP analysis |
杨树溃疡病菌是造成我国北方杨树死亡的重要原因之一(向玉英,1987)。该菌致病力的分化早有报道,但分化的细胞学基础尚未有人研究。本文研究了病菌营养体自然条件下亲和得到的3株异核体菌株,并从细胞学的基础上证明异核体菌株确实为病菌营养体亲和的结果。通过接种实验和统计分析,研究了异核体菌株与亲本菌株致病力差异,异核体菌株致病力明显低于亲本菌株致病力,异核体菌株间致病性存在显著分化。
本研究表明:异核体现象是造成溃疡病菌致病力分化的重要原因之一。异核体形成并不产生明显的异核体优势,异核体菌株致病力与弱毒亲本菌株毒力相近,而与毒力强的亲本毒力差异显著。核糖体ITS区段是研究真菌系统发育和群体遗传变异的有效片段(余仲东等,2000),对异核体菌株与亲本菌株ITS-rDNA-RFLP分析表明,异核体菌株和亲本菌株并无显著核遗传差异,说明在菌丝融合过程中,核质因素并不是菌株致病性差异的主要原因,弱毒株系胞质流动可能是造成毒性变异的重要因素。在板栗疫病的研究中也发现弱毒株系细胞质在营养体亲和群体(VCGs)中流动,尤其是其中dsRNA的传递是造成菌株弱毒的原因(王克荣等,1997)。本研究中2亲本菌株致病力并无统计上的显著差异,但接种结果表明,弱毒亲本是影响异核体毒力显著弱化的原因。异核体菌株(AB)具有明显的双亲本生长特性,也能正常形成分生孢子,但尝未发现子囊壳形成。少部分分生孢子菌落在培养中发生回复突变现象,表现出A或B菌株培养特性,更多的像A菌株,大部分分生孢子菌落在连续转接培养中表现出稳定的培养生物学特性。这种现象说明,异核体菌株中细胞核仍存在重组分化,其机理和作用尝不清楚。
Fulgen染色法是将核酸中的五炭糖氧化而显色,在植物细胞学研究中应用较多。本研究表明该法也是真菌细胞学研究的良好方法。同Gimsa染色法一样,该法对菌丝体隔膜区分上存在困难(罗信昌,1998),尤其是对纤细的气生菌丝。在供试菌株ITS-rDNA-RFLP分析中,本研究只使用了2种高频限制性内切酶,该菌是否存在其他位点的变异,值得进一步研究。
康振生, 李振岐. 1993. 小麦条锈菌异核作用产生的一新菌系. 西北农业大学学报, 21(1): 97-99. DOI:10.3321/j.issn:1671-9387.1993.01.020 |
罗信昌. 1998. 木耳和毛木耳的极性研究. 真菌学报, 7(1): 56-61. |
王克荣, 成桂英, 刘怡君, 等. 1997. 栗疫病菌的营养体亲和性基因和对dsRAN病毒传播的影响. 菌物学报, 16(1): 30-35. |
向玉英. 1987. 杨树病害及其防治. 北京: 中国林业出版社.
|
余仲东, 张星耀, 曹支敏. 2000. 真菌核糖体基因间隔区研究概况. 西北林学院学报, 15(2): 107-111. DOI:10.3969/j.issn.1001-7461.2000.02.023 |
余仲东, 张刚龙, 曹支敏. 2002. 病原菌种内群体遗传分化研究现状与技术. 西北林学院学报, 17(4): 66-72. DOI:10.3969/j.issn.1001-7461.2002.04.018 |
郑若立. 1978. 实用细胞学技术. 北京: 科学出版社.
|
Zhu H, Qu F, Zhu L H. 1993. Isolation of genomic DNAs from plants, fungi and bacteria using benzyl chloride. Nucleic Acids Research, 21(22): 5279-5280. DOI:10.1093/nar/21.22.5279 |