 2006, Vol. 42
2006, Vol. 42文章信息
- 周成刚, 张卫光, 乔鲁芹, 孙绪艮, 王志勇.
- Zhou Chenggang, Zhang Weiguang, Qiao Luqin, Sun Xugen, Wang Zhiyong.
- 东方真叶螨的生物学特性、有效积温及发生规律
- The Biological Characteristics, Effective Accumulate Temperature and Occurrence Regulation of Eutetranychus orientalis
- 林业科学, 2006, 42(5): 89-93.
- Scientia Silvae Sinicae, 2006, 42(5): 89-93.
-
文章历史
- 收稿日期:2005-03-01
-
作者相关文章



2. 泰安市泰山风景名胜区管理委员会 泰安 271018
2. Management Committee of Mountain Tai'an Famous Scenery Tai'an 271000
东方真叶螨(Eutetranychus orientalis)在山东泰安、济南、莱芜等地是大叶黄杨(Euonymus japonicus)的重要害螨(王慧芙,1981)。近年来,由于该螨的危害,造成大叶黄杨叶片早期枯黄脱落,严重影响了其观赏、绿化价值。据在泰安市调查,行道两侧的大叶黄杨绿篱和绿球受害率极高,有的路段可达100%。大叶黄杨受害后,叶绿素含量下降,光合速率降低,细胞间隙CO2浓度增高,气孔导度降低,蒸腾速率下降,生长量降低,受害越重,影响越大。因该螨体型微小,繁殖速度快,不易发现,其危害有越来越重的趋势。国内对该螨的研究报道甚少,仅见有王慧芙(1981)、忻介六(1988)对其形态特征的记述。林延谋等(1995)报道了该螨在橡胶树(Hevea brasiliensis)上的繁殖和发育;孙绪艮等(1996a)研究了该螨对寄主植物的危害性,其他尚未见报道。本文研究了该螨在山东大叶黄杨上的生物学特性和有效积温,以期为防治该螨提供依据。
1 材料与方法 1.1 生物学供试叶螨采自山东农大校园内的大叶黄杨。室内用海绵水盘法(孙绪艮等,1996a)饲养,将室外采回的雌成螨用小毛笔接于叶片上,置于(25.0±0.5) ℃的全光照培养箱内,待其产卵24 h,每盘内有卵50粒以上时除去雌成螨。再分别将卵放入20.0、22.5、25.0、27.5、30.0 ℃(温差±0.5 ℃)全光照条件下的生化培养箱内,记录卵的历期。幼螨孵出后,转移至同样条件下的海绵水盘内单头饲养,直至成螨死亡为止,记录幼螨至成螨历期。每温度处理重复3次。在以上研究的同时,观察其生长发育历期、蜕皮、交尾、产卵等习性。
1.2 有效积温将1.1饲养所得的各螨态历期,用以下4种方法计算各螨态及1个世代的发育起点温度和有效积温。
1.2.1 最小二乘方法根据有效积温法则K=N(T-C),将试验数据代入公式得C、K以及各发育阶段发育起点温度C的标准误差Sc.
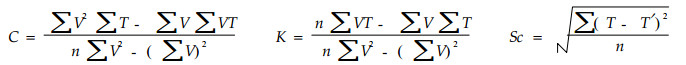
|
式中,C为发育起点温度,K为有效积温,V为发育速率,T为平均温度,n为处理数,Sc为发育起点温度的标准差,T′为理论温度。
1.2.2 直线回归法根据有效积温法则的第2种表达形式T=C+KV, 将试验数据所得的T和V,用直线回归法求出C、K,并求出相关系数。
1.2.3 最小变异系数法根据最小二乘方法求得的发育起点温度,假设卵的若干个C,分别为6.0,6.5,7.0,7.5,8.0,8.5,9.0 ℃,幼螨至成螨的若干个C分别为4.5,5.0,5.5,6.0,6.5,7.0,7.5 ℃,全世代的若干个C分别为5.0,5.5,6.0,6.5,7.0,7.5,8.0 ℃,按以下公式求得变异系数c.v.
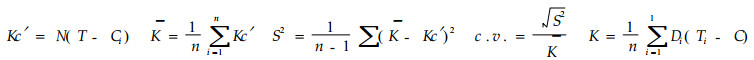
|
式中,Kc′为理论有效积温,K为平均有效积温,N为发育历期,s2为方差。
求得变异系数c.v.最小的所对应的温度即为其发育起点温度。然后求得各发育阶段的有效积温K,并据1.2.1中的公式求出Sc。
1.2.4 直接最优法用极值理论建立目标函数O.F.,当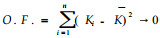
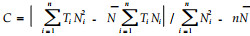
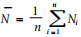
根据K=N(T-C), 将以上4种方法求得的发育起点温度C和有效积温K,代入不同温度,求得卵期的预测值,与实际历期比较验证。于3月份寻找越冬螨,待叶片上开始出现活动螨后,定期观察记载其活动情况,直到叶片上不再有螨活动为止。应用4种方法求得的C值、K值和泰安市气象资料,计算东方真叶螨在泰安的理论年发生代数,并与2000年室内单头饲养结果比较。
1.3 发生规律与种群消长 1.3.1 越冬螨态及越冬部位2002年3月在室外选定15个大叶黄杨枝条,去除尘土后,缠两层双面胶带,做好标记,逐日记录双面胶带上是否粘有东方真叶螨成螨。并调查下部50叶片,检查是否有越冬卵。此外,在室内盆栽大叶黄杨上接种饲养各螨态若干,10月初在枝条下部缠双面胶带,观察是否有雌成螨下树越冬。
1.3.2 越冬卵孵化时期及进度从室外采集带有越冬卵的叶片,剔除空卵壳、死卵及杂物等,留活卵300粒以上,自4月上旬开始用海绵水盘法逐日观察孵化数,至孵化结束。
1.3.3 年间种群消长规律4月中旬开始,在大叶黄杨绿篱上分上、中、下层随机采集大叶黄杨叶片150个,立体显微镜下观察记录各螨态数量,每10天左右调查1次,直至叶片上无活动螨为止。
2 结果与分析 2.1 生长发育单头饲养结果表明,东方真叶螨的全部发育过程为:卵→幼螨→第1若螨→第2若螨→成螨。在幼螨至成螨的形态转变之前,均有1个静止期。雌、雄个体均经过第2若螨期。在5种不同温度下饲养,各发育阶段的历期如表 1所示。在20~30 ℃范围内,各发育阶段的历期随温度的升高而缩短。
|
|
如2.1所述, 东方真叶螨在幼螨至成螨的发育过程中,有3个静止期。静止时,足卷缩于体下,不食不动,身体紧贴于叶面上,静止结束后脱皮变为下一螨态。蜕皮时表皮在第2、3对足之间作横向裂开,先蜕下后半部皮,体向后缩,再把前半部皮脱下。
2.2.2 交配雌螨一经蜕皮变为成螨后,随即与提前羽化的雄成螨交配,交配时,雄螨爬至雌螨腹面,腹部末端上弯,阳具伸入雌性生殖孔内进行交配。一次交配时间长短不一,长时持续数分钟,短时不足1 min。雌、雄成螨均可多次交配。
2.2.3 产卵与成螨寿命雌成螨羽化后1~2 d开始产卵,产卵历期6~8 d,第3~5 d达到高峰。单雌产卵量25粒左右。雌成螨产卵后在卵周围不停活动,以口器吐细丝将卵连接于叶面上,颚体在卵边缘和叶面相接处不住摆动,便有一层膜形成,将卵封固,具明显的“护卵”习性,前后约需5~8 min。但无吐丝结网习性。在22.5~27.5 ℃范围内,雌成螨寿命8~10 d,雄成螨寿命4~6 d,并随温度的升高而缩短。
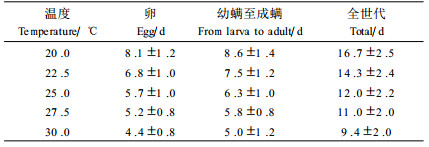
2.3 有效积温对有效积温的推算,各学者采用不同的计算方法,为明确各方法的计算结果是否一致,本文用4种方法计算比较有效积温和发育起点温度,结果如表 2所示。不同计算方法所得结果有所不同,但差异较小。如幼螨至成螨和全世代的发育起点(C)最多相差不到0.5 ℃;卵期(C)相差0.8 ℃。有效积温(K)相差4.2~4.4 d ℃。差异均达不到显著水平。表明用这几种方法计算所得发育起点和有效积温均可靠,无显著差异。
|
|
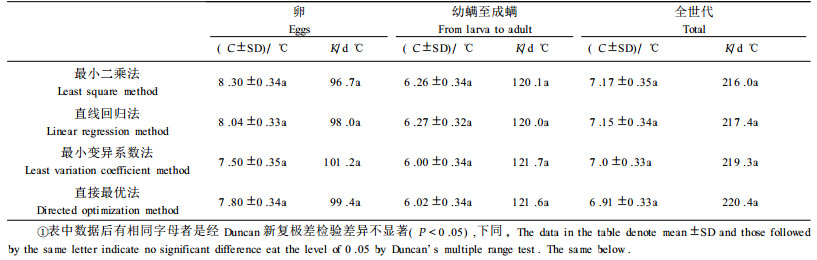
以卵历期为例,验证不同计算方法求得的发育起点温度C值的可靠性。结果如表 3所示。各计算方法所得的预测值与实际值,均无显著性差异。因此,在实际运用过程中,可选用较简单的方法,笔者认为,直线回归法具有快速、简便的优点,较为实用。
|
|
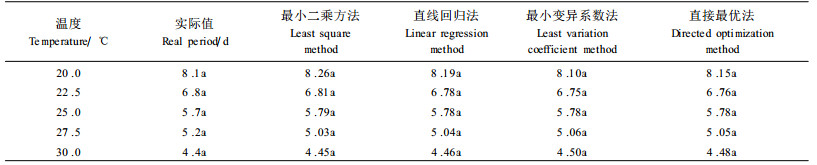
根据泰安地区气象资料,及东方真叶螨全世代的发育起点温度和有效积温,求得总有效积温K1,根据发生代数=K1/K,将预测结果列入表 4。结果说明,四种方法的预测结果差别不大,均在13~14代之间,与2000年室内单头饲养代数(13代)基本一致。
|
|

叶螨多以雌成螨在翘皮裂缝、枯枝落叶等隐蔽场所越冬,或是以卵在枝干上越冬(忻介六,1988)。室外调查大叶黄杨50个下部叶片,其上有越冬卵26粒,平均密度达每页0.52粒,由此证明东方真叶螨在室外不以雌成螨越冬,而以卵在大叶黄杨下部叶片上越冬。但室内调查盆栽苗上的发生情况,发现有雌成螨下树(表 5),同时叶片上仍有幼、若螨。分析认为:室内温度较高,能够生长发育、繁殖,随着温度下降,叶片逐渐老化,不能满足其继续生长发育的需要,雌成螨开始下树潜藏越冬,据此推测,该螨在气温较高的温暖地区可能以雌成螨在树下隐蔽场所越冬。
|
|
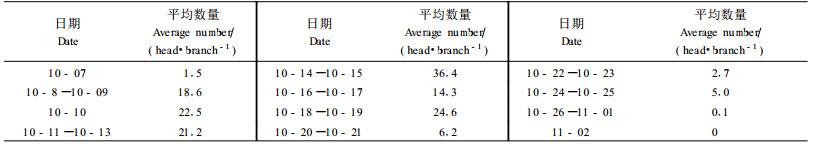
如图 1所示,越冬卵4月13日至14日开始孵化,孵化盛期在4月15日至4月20日,4月26日孵化结束。该期间正值大叶黄杨刚开始旺盛生长期,因为越冬卵基数较少,第一代螨口密度较低,一般不会对生长造成大的影响。但随着气温的增高,繁殖速度的加快,危害也渐趋严重。

|
图 1 东方真叶螨越冬卵孵化进度 Fig. 1 Hatching progress of overwintering eggs of Eutetranychus orientalis |
东方真叶螨自4月中旬至10月中旬各螨态均有发生(图 2)。其发生全年有2个高峰:第1个在5月下旬至6月中旬,第2个在9月中、下旬,以第1个高峰期螨口密度大,危害严重,短期内可导致大叶黄杨叶片变色。6月下旬至8月下旬螨口密度很低,主要原因是:因第1个高峰期危害严重,很快造成叶片营养恶化,不能全面满足东方真叶螨的营养需要,致使其死亡率高,加上该期进入雨季,由于雨水的机械冲刷作用,从而造成螨口密度低,此外,该时期气温较高,不适于其生长发育,也是造成其死亡率高的原因。在调查中发现,东方真叶螨在早期有从下层逐渐上移危害的习性,该时期修剪大叶黄杨能压低螨口密度。

|
图 2 东方真叶螨年间种群消长曲线 Fig. 2 Eat and flow curve of Eutetranychus orientalis population |
生长发育 本研究发现:东方真叶螨的雄性个体不缺少第2若螨期。忻介六(1988)指出,有些叶螨的雄性个体在发育过程中,缺少第2若螨期。但孙绪艮(1992)曾观察过5种叶螨,山楂叶螨(Tetranychus viennensis)、朱砂叶螨(T. Cinnabarinus)、柑桔全爪螨(Panonychus citri)、苹果全爪螨(P. ulmi)、针叶小爪螨(Oligonychus ununguis),以及孙绪艮等(1995;1997)对桑始叶螨(Eotetranychus suginamensis)及竹裂爪螨(Schizotetranychus bambusae)的研究中均未发现此种现象,仅在杨始叶螨(Eotetranychus populi)发现极个别出现无第2若螨的现象(孙绪艮等,1996b),其余均有第2若螨,出现的原因尚不清楚。笔者在对东方真叶螨的研究过程中,在不同的温度处理下所有个体均存在第2若螨期。
有效积温 对同一种节肢动物来讲,各种方法计算出的发育起点温度低,则其有效积温就高,这在以往的研究和本研究中都得到了证实。但这是在实验室恒温条件下得到的结论,在自然变温情况下如何,尚需验证。
年间种群消长规律 影响东方真叶螨年间种群消长规律的因子很多,不同年份环境变化特别是降水变化会较大,而降水能显著影响其种群数量。因此,本文中仅调查1年的数据是有限的,还应作更多年份的统计调查,结合气象资料,才能说明其年间种群消长规律及其与气候条件的关系。
林延谋, 符悦冠, 杨光融, 等. 1995. 温度对东方真叶螨的发育与繁殖的影响. 热带作物学报, 16(1): 94-98. |
孙绪艮, 刘振宇, 周成刚, 等. 1996a. 东方真叶螨对大叶黄杨的危害. 植物保护, 22(5): 29-30. |
孙绪艮, 周成刚, 刘玉美, 等. 1996b. 杨始叶螨生物学和有效积温研究. 昆虫学报, 39(2): 166-171. |
孙绪艮. 1992. 五种叶螨个体发育观察. 昆虫知识, 29(5): 277-278. |
孙绪艮, 周成刚, 刘玉美, 等. 1995. 桑始叶螨生物学特性研究. 蚕业科学, 21(3): 184-185. |
孙绪艮, 周成刚, 刘玉美, 等. 1997. 竹裂爪螨生物学特性的初步研究. 林业科学, 33(3): 274-277. DOI:10.3321/j.issn:1001-7488.1997.03.012 |
王慧芙. 1981. 中国经济昆虫志:二十三册.叶螨总科. 北京: 科学出版社, 43-44.
|
忻介六. 1988. 农业螨类学. 北京: 农业出版社, 59.
|