 2006, Vol. 42
2006, Vol. 42文章信息
- 郭泺, 夏北成, 李楠, 余世孝.
- Guo Luo, Xia Beicheng, Li Nan, Yu Shixiao.
- 快速城市化过程中深圳森林小群落结构特征及其多样性研究
- Analysis of Microcommunity Structure and Its Diversity of Shenzhen Urban Forest during Rapid Urbanization
- 林业科学, 2006, 42(5): 68-74.
- Scientia Silvae Sinicae, 2006, 42(5): 68-74.
-
文章历史
- 收稿日期:2005-02-28
-
作者相关文章



2. 深圳市仙湖植物园 深圳 518004
2. Fairylake Botanical Garden Shenzhen 518004
区域城市化是城市扩展的影响过程,其实质是区域自然生态系统和农业生态系统向城市生态系统不断转化的过程(宋治清,2004)。这一过程在快速城市化条件下导致城市景观结构剧烈变化,一个基本特征是城市森林自然植被覆盖面积减少,这暗示着地表生态环境容量下降(曾辉,2000;周志翔,2004)。林地的转移、道路的切割、旅游业的侵占以及城市工业园地及居民和特殊用地的扩张,使连片集中的结构上具有完整性、相对独立的大片森林植被分割转变成小块状、不连续斑块或片状的“城市森林”(朱文泉,2003;胡志斌,2003)。由于城市土地较高的商品价值,因此有人提出城市不应有森林的观点,对城市进行美化绿化即可,这给城市森林的存在带来了极大的压力。在城市森林缀块生境上残存的森林,实质上已形成为特殊的森林小群落。这些小群落,是城市最为宝贵的自然资源,在城市区域自然生态系统中最具生态学意义。它不同于人工植被,它的存在和保护虽然受到了高度重视和关注,但现实表明城市森林还是在不断衰退。城市森林小群落的衰退及其生境上的脆弱和不稳定性对城市生态安全的潜在影响远比资源减少要严重得多。城市森林小群落的衰退、消失,尤其是结构和物种多样性的特征变化所带来的城市生态环境安全问题比绿地植被面积减少,更为荫蔽和严重,在城市化过程中给予应有的关注,意义十分重要。
在城市区域内森林植被各种群落空间复合体可以理解为小群落复合体系,其缀块生境虽然相隔一定距离,但具有同一系统的特征,其整体作用可影响区域内一定范围的外界环境,同时也受制于环境。从而形成独特的缀块生境和不同的群聚群落,其中生态适应和自然选择起着决定性作用。小群落的各种特征,包括数量特征、丰富度、均匀度特征和物种多样性等,都包含丰富而重要的生态学内容(Loetsch,1973;王伯荪,1978;Howlett et al., 2000;Chapin et al., 2000;蒋有绪,2002)。与一般生态系统一样,其中起着控制作用的是那些小群落中的优势种。优势种既是一个群落或一个种群空间复合体的骨架,又决定其外貌和结构,因此研究城市区域小群落的优势种是其前提和基础。本文以深圳市为研究区域,引用岛屿生物地理学和复合种群理论,将市域内遗留的“森林”(不包括人工植被和绿化系统)缀块生境比照为岛屿状生境,选择典型小群落,综合描述和比较深圳市森林小群落多样性状况以探讨内在的生态学意义。
1 研究区与研究方法 1.1 自然概况深圳市是中国南部沿海城市,位于北回归线以南,113°46′—114°37′E,22°27′—22°52′N,东临大亚湾和大鹏湾,西濒珠江口和伶仃洋,南边深圳河与香港相连。亚热带海洋性气候,气候温和。全境地势东南高,西北低。大部分为低山、平缓台地和阶地丘陵。深圳市是我国第1个经济特区,经过20年由一个3万人口边陲小镇飞跃发展成为特大城市,完成了从传统农业地区向新兴的高度城市化地区转化过程,城市化水平的快速提高,土地利用和覆盖的变化带来深刻的生态环境影响,土地覆盖格局变化表现为由特区内向特区外梯度转变的空间分异规律。地带性植被类型为热带季雨林型的常绿季雨林,组成种类多样,天然植被有季雨林,常绿阔叶林、红树林、竹林、灌丛和草丛等,其原生植被早已不复存在,遗留的“森林”符合小群落的界定,均生存在充满缀块性的或破碎化的城市景观中,林地在空间上是孤岛式的聚集分布。
1.2 研究方法在深圳市的笔架山、小南山、三洲田、梧桐山、莲花山、下围岭等6处“遗留的森林生境”林地上,分别以乔木层优势树种为标准,设置以10 m×10 m样方组合的样地,包括200、400、1 200 m2 3种尺度的取样面积,利用GPS定位,导入深圳数字地形图,坐标配准,共计设样方48块(图 1)。采用常规每木调查法,主要测定胸径在1.0 cm以上、高度在1.5 m以上各树种胸径(D)、树高(H)。采用群落学统计方法主要统计树种个体数(N)、树种数(S)、单个体树种数(S1)、双个体树种数(S2)以及平均直径(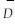
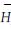
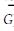

|
图 1 样地数字分布图 Fig. 1 Distribution map of plot sites in the study region |
Simpson多样性指数: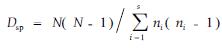
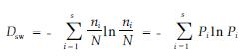
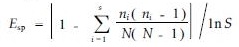

为描述方便,本文提出蓄积水平级概念,即种群每公顷蓄积量的水平等级。蓄积量采用平均试验形数法求积式求算,并以其整数值定名为蓄积指数(Vg)
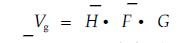
|
式中: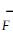
所研究的森林群落平均密度为0.689株·m-2;优势种的总个体数占55.3%,平均密度为0.276株·m-2,平均直径10.0 cm,平均高8.8 m,平均蓄积指数49,总体蓄积水平为Ⅱ级。从表 1可知Ⅰ级水平的优势种为桉树(Eucalyptus globulus),其蓄积量大于平均值的80%,桉树株数占群落总株数的54.4%。且蓄积指数值大。该桉树林物种数和单个体数均较少,林内乔木层只有少量荔枝(Litchi chinensis)伴生,其余为野桐(Mallotus japonicus)、楠木(Phoebe zhennana)、九节(Psychotria rubra)、马樱丹(Lantana camara)等幼苗。林内未发现桉树幼苗生长,表明天然更新不良,种群一旦被损,九节等将替代。
|
|
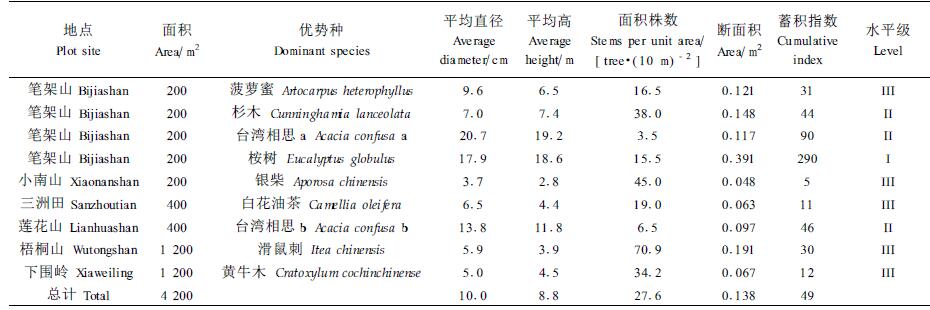
Ⅱ级水平的优势种群有杉木(Cunninghamia lanceolata)和台湾相思(Acacia confusa)。杉木种群在群落中处于绝对优势地位。杉木种群的平均直径和平均高虽低于总平均值,但株数和断面积G均大于总平均值。乔木层中有桉树、马占相思(Acacia mangium)、花椒(Zanthoxylum bungeanum)等伴生,表明林内种间竞争将会更趋复杂激烈。林内幼苗和小树比重较大,分别占42.5%和41.9%,其中杉木的幼苗和小树就分别占到11.7%和20%,说明林内更新良好。台湾相思在群落中虽居于林冠上层并形成单层纯林结构,但面积株数和断面积水平较低,正处于衰退状态,林内已被九节、梅叶冬青(秤星树)(Ilex asprella)、银柴(Aporosa chinensis)、山油柑(降真香)(Acronychia pedunculata)等物种占据,幼树和小树株数占有相当大的比重,分别占52.7%和39.2%。
Ⅲ级水平的优势种群有波罗蜜(Artocarpus heterophyllus)、银柴、油茶(Camellia oleifera)、滑鼠刺(Itea chinensis)和黄牛木(Cratoxylum cochinchinense)。乔木层有梓树(Catalpa ovata)、NFDA3档(画眉跳山胡椒,鹰不泊)(Zanthoxylum avicennae)、火灰山矾(Symplocos dung)、马尾松(Pinus massoniana)、梨润楠(Machilus pomifera)、棱枝冬青(Ilex angulata)、鹅掌柴(鸭脚木)(Schefflera octophylla)、乌材(Diospyros eriantha)、尖峰润楠(Machilus monticola)等多物种占据树冠上层,但层次结构已经不明显,各优势种群总体上平均直径、平均高和断面积均低于总平均值,蓄积指数均低于40,其主要特征是林内混生物种多,并且多为小树。
综观各优势种群所处的群落,具有层次分化不明显,径级和高度偏低的特点,说明各群落均处于群落演替过程之中,丰富的多物种幼苗和小树特征指示了阳生性树种将逐渐取代而成为优势种。
2.2 群落结构特征 2.2.1 物种个体数的直径分布物种个体数明显地与直径呈近似双曲线型的反J形分布,随直径径阶增大,个体数减少的趋势十分明显,这与寇文正(1982)的研究相吻合,可采用β分布拟合。本文采用幂回归式y=A·Xβ,拟合效果亦较理想。经验式为:N=17 028.61D-2.716 7 (r=0.952)。
图 2说明了物种个体数主要集中在小径阶的2~6 cm直径级范围内,个体数随直径增大,到2~3 cm径阶时急剧减少的趋势非常明显,个体数减少率达32.5%;3~6 cm径阶仍表现为急剧下降特征,个体数减少率为15.3%,6 cm径阶之后,曲线渐趋平缓,个体数减少的幅度逐渐变小。为此可将6 cm径阶作为物种个体数直径分布变化分界点。各群落优势种的个体数直径分布,具有与9个种群物种个体数直径分布相同规律。

|
图 2 物种个体数的直径分布 Fig. 2 Diameter distribution of number of individuals for species |
图 3说明了波罗蜜、杉木、银柴、油茶、台湾相思5个群落优势种个体数的直径分布。趋势与图 2所示结果一致,表明当优势种个体数大并具有一定的直径分布结构时,优势种的个体数直径分布可以反映群落个体数直径分布规律;当优势种个体数偏小且直径分布零散时,如笔架山样方的台湾相思和桉树2个优势种,就难以反映这种分布规律。

|
图 3 5个群落优势种个体数的直径分布 Fig. 3 Diameter distribution of individual number for 5 dominant species in communities |
上述规律表明在物种多样性分析时不能单纯依赖物种个体数,还应考虑到群落个体数的直径结构。在2 cm直径级内,幼苗大量聚集必然产生较多的物种数,稀有种和偶见种或单个个体种、双个体物种出现的机率就高;在3~5 cm直径级内,小树大量聚集,受竞争密度效应影响物种消失的可能性亦将增大;6 cm以上直径级的林木个体,生长比较稳定,物种数的变动较稳定,这都与个体数的直径结构有关。研究中是包括幼树的全部个体数(D≥1.5 cm),还是不包括幼树的个体数(D≥2.5 cm),或只包括大树的个体数(D≥5.5 cm),这需要进行具体分析。已有的一些多样性指数只是依赖于物种个体数,并未考虑种间个体数的差异,更未考虑个体数的直径分布结构,显然是不合理的,因此很难保证可比性。
2.2.2 个体数的高度分布规律研究区群落物种个体数85.9%集体分布在1.5~4.5 m高度级区间范围内,且59.0%集中分布在4.5 m高度级中,说明个体数高度普遍偏低且分布十分集中。不同群落的个体数高度级分布规律同样表现为随高度级增高,个体数波动性减少趋势(图 4)。这种波动趋势在1.5~4.5 m高度级区间表现出与取样面积有关,A=200 m2的01~05号群落呈下降趋势;A=400 m2以上的群落则呈上升趋势,且取样面积越大在1.5~4.5 m高度级区间,上升趋势越明显。进一步分析200、400和1 200 m2在1.5~4.5 m高度级区间内个体百分数分别为24.7%、69.5%和88.9%,说明尽管具有这种波动,取样面积的大小并不影响个体数在这一高度级区间内的比例。

|
图 4 群落个体数高度级分布 Fig. 4 Height distribution of individual number of communities |
不同群落占据有不同高度级的空间范围,其物种个体数的变化亦很大,这与群落蓄积水平级相关。在1.5~4.5 m区间Ⅰ、Ⅱ、Ⅲ蓄积水平级平均个体数依次占42.1%、69.0%和89.7%;在7.5~13.5 m区间依次占5.3%、27.2%和10.3%;在16.5~25.5 m区间依次占52.6%、5.8%和0。可以看出Ⅰ蓄积水平的群落个体主要占据了最高高度级区间。Ⅱ蓄积水平的群落个体主要占据了中等高度级区间,而Ⅲ蓄积水平的群落个体则主要占据低高度级区间。
群落个体数的高度结构变化规律说明在物种多样性分析时还应考虑个体数的高度结构。不同群落或相同群落的平均高度不同,物种个体数的变异亦很大,所占据的高度级区间范围亦有区别,也影响物种多样性测度结果和可比性。森林群落树高曲线一般为抛物线形,物种的高度随直径的增加而增加,个体数随直径的增加而减少,这使平均高度这一指标在个体数高度分布格局中占有重要地位。由于平均高是蓄积量计算的重要因子,当形数一定时,物种的重要性仅考虑断面积因子显然欠妥,因此在物种多样性分析时,还应考虑高度问题,群落物种个体数在不同高度级区间的比例和分布格局,值得森林植物多样性研究关注。
2.2.3 个体数与物种数关系分析3种起测径阶的物种个体数(N)与物种数(S)之间呈抛物线形关系,经采用y=a+blnX式拟合,效果极为理想,r值均在0.91以上。不同的起测径阶在相同N条件下所表征的S值大小及其变化规律为D1>D2>D3,起测径愈小,N愈大,每增加100个N数时,S的增值也愈大,表明物种数的多少与量测时所确定的起测径阶大小有关。起测径阶越小,在相同个体数条件下所反映的物种数越多。由表 2可推算出不同的个体规格、不同个体数时的物种数。表明无论是幼树、小树或大树,物种数均随物种个体数的增加而增加,不同个体规格的物种总个数在20~160范围内时,物种数增加率在2.4%~5.9%之间,总增加率为11.2%, 即每增加100个个体数,可增加11.2个物种。
|
|

在取样面积A≤1 200 m2条件下,个体数与物种数的抛物线曲线关系表明了S随N而增加的变化趋势,其变化率与起测径阶大小有关,实际上还受更多因素的影响,如面积大小、物种种类以及缀块生境等因素的综合影响。N与S的关系是复杂的,在构建物种多样性表述模型时,应尽可能多地考虑不同情况下的N与S关系,在取样面积一定时(A≤1 200 m2),群落中幼树和小树的总个体数占69.4%,个体数与物种数之间的变化增加率占47.3%,与大树52.7%较接近,因此在物种多样性分析时,考虑层次结构上的差异亦十分必要。
2.3 群落多样性分析物种多样性与物种数和个体总数有关,还与物种的均匀度有关(彭少麟,1996)。本文对多样性指数特征从丰富度、均匀度和多样性3个方面进行分析,并研究了其各指数值的相关性(表 3)。
|
|
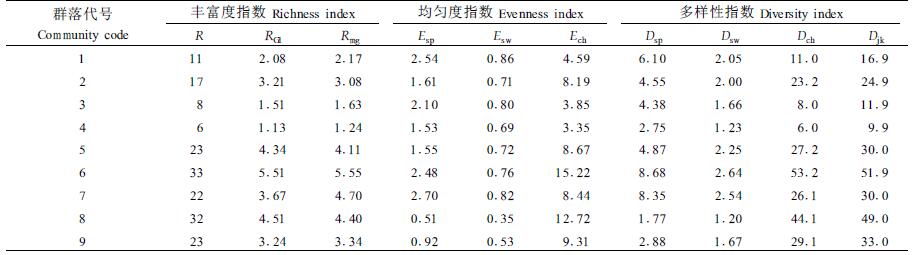
选用了3种丰富度指数,其中R指数只涉及到群落物种数本身,是个直观的指数值,为便于各群落比较,取样面积400和1 200 m2时R的值均分别以0.072 5和0.024 5的调整系数调整到200 m2取样面积上;9个群落R值范围在6~33之间;可见R值表达丰富度虽然直观,但变异较大,RG1指数涉及到S与A的关系,Rmg指数涉及到S与N的关系,二者指数值分别在1.13~5.51和1.24~5.55之间。但RG1和Rmg指数并未考虑种间个体数的差异,可能忽略常见种(common species)和稀有种(rare species)对群落多样性贡献的差异,3种丰富度指数所表达的指数值在各群落中排序基本相似。
2.3.2 均匀度指数特征选用Esp、Esw和Ech3种均匀度指数,其计算涉及到Dsp、Dsw和Dch 3种不同的多样性指数与S的关系。Ech均匀度指数值较大,群落变化范围在3.35~15.22之间,虽然未考虑种间个体数差异,但考虑到了群落中单个体S1与双个体数S2之间的关系,保证和体现了稀有种和偶见种在群落多样性中的突出贡献;Esp和Esw均匀度指数值相比较Ech值小,各群落变化范围分别在1.53~2.70和0.35~0.86之间。由于3种均匀度指数所表达的指数值在研究群落中排序差异较大,但又各具特色,因此,那一种均匀度指数更能反映物种均匀程度,尚需比较和探讨。
2.3.3 多样性指数特征选用了4种多样性指数,Dsp多样性指数采用了Simpson多样性表达式作为测度指标以反映其物种的多样性,各群落的指数值变化范围为1.77~8.68;多样性指标Dsw与Dsp性质相同,同样考虑到了物种个体数与物种总个体数之间的比例关系而较为普遍地被采用,各群落的指数值变化范围在1.20~2.64之间。由于Dsp与Dsw同样考虑到了种间个体数在群落中的比例差异因素,保证和体现了常见种、稀有种、偶见种在群落多样性中的微妙贡献,因此在群落指数值排序上表现出相似性,Dck和Djk多样性指数是建立在物种数、单个体物种数、双个体物种数之间及与总个体数之间关系的多样性指数。是考虑到热带雨林里一定面积样地中相当多的物种仅有1、2个个体特点,而在近些年才提出的非参数指数,即Chao多样性指数和二阶刀切法多样性指数(余世孝等,2001)。各群落的Dch和Djk指数值变化范围分别为6.0~53.2和9.9~51.9之间。由于Dch和Djk都同样考虑了单个体数与双个体数在多样性中的贡献,突出了罕见种或偶见种在维持群落物种多样性的作用,因此各群落的指数值排序上,表现为较高的排序吻合率。
2.3.4 物种多样性综合描述分别对各群落的各种丰富度指数、均匀度指数和多样性指数计算平均指数值和平均指数综合值K(简称综合指数K)(表 4)。综合指数K值的大小,实际上反映了群落物种丰富度、均匀度和多样性的平均水平,K值百分比则表达了群落物种的综合指数K占全部群落的比重,K值的百分比大小排序表达了各群落物种综合多样性比较的结果。
|
|

三洲田样方以油茶为优势种的群落物种综合指数K值最大为16.65,排序第1,其他依次为梧桐山的滑鼠刺、莲花山的台湾相思、小南山的银柴、下围岭的黄牛木为优势种的群落,K值在9.80~14.06之间;笔架山的杉木、波罗蜜、台湾相思和桉树为优势种的各群落,K值相对小,依次为8.31、5.58、4.07和3.21。各群落平均丰富度指数8.67,平均均匀度指数3.55,平均多样性指数15.11,平均综合指数K为9.11。
3 结论和讨论城市森林小群落多样性分析不能单纯依赖物种个体数,还应考虑个体数的直径级和高度级结构分布规律,否则将影响物种多样性测度结果和可比性。基于群落组成和特征结构的综合性物种多样性分析方法,将受到更多关注。采用丰富度、均匀度、多样性以及复杂性等各种指数相结合的综合指数及排序方法,可用于群落多样性比较。为提高可比性,应统一测度标准,包括测度起测径阶、直径级和高度级区间、取样面积,对于指数的选择有待进一步探讨。
深圳城市森林小群落总体上具有直径和高度偏低、个体数相对集中、层次分化不明显和各群落均处于群落演替过程的特点。丰富度平均指数在2.79~14.69之间,均匀度平均指数在1.86~6.15之间,物种多样性平均指数在4.97~29.11之间,综合指数K值在3.21~16.65之间。小群落在取样面积为200 m2的条件下,平均综合指数K值为9.11。平均综合指数从大至小排序结果为:三洲田油茶>梧桐山滑鼠刺>莲花山台湾相思>小南山银柴>下围岭黄牛木>笔架山杉木>笔架山波罗蜜>笔架山台湾相思>笔架山桉树。
胡志斌. 2003. 沈阳市城市森林结构与效益分析. 应用生态学报, 14(12): 2108-2112. DOI:10.3321/j.issn:1001-9332.2003.12.005 |
蒋有绪. 2002. 海南岛热学林生物多样性及其形成机制. 北京: 科学出版社, 25-68.
|
寇文正. 1982. 林木直径分布的研究. 南京林业大学学报, (2): 51-56. |
彭少麟. 1996. 南亚热带森林群落动态学. 北京: 科学出版社, 31-42.
|
宋治清. 2004. 城市景观及其格局的生态效应研究进展. 地理科学进展, 23(2): 162-165. |
王伯荪. 1978. 植物群落学. 北京: 高等教育出版社, 65-83.
|
余世孝, 臧润国, 蒋有绪. 2001. 海南岛霸王岭垂直带热带植被物种多样性的空间分析. 生态学报, 21(9): 1438-1443. DOI:10.3321/j.issn:1000-0933.2001.09.006 |
张金屯. 1994. 植被数量生态学方法. 北京: 中国科学技术出版社, 15-35.
|
曾辉. 2000. 深圳市龙华地区快速城市化过程中的景观结构研究——林地的结构和异质性特征分析. 生态学报, 20(3): 378-383. |
周志翔. 2004. 城市绿地空间格局及其环境效应. 生态学报, 24(2): 186-192. |
朱文泉. 2003. 城市森林结构的量化研究——以沈阳树木园森林群落为例. 应用生态学报, 14(12): 2090-2094. DOI:10.3321/j.issn:1001-9332.2003.12.002 |
Chapin F S Ⅲ, Zavaleta E S, Eviner V T, et al. 2000. Consequences of changing biodiversity. Nature, 405: 234-242. DOI:10.1038/35012241 |
Connell J. 1978. Diversity in tropical rainforests and coral reefs. Science, 199: 1302-1310. DOI:10.1126/science.199.4335.1302 |
Howlett, Dhand R. 2000. Nature insight biodiversity. Nature, 405: 207. DOI:10.1038/35012215 |
Loetsch F.1973.Forest Inventory, Vol. Ⅱ Mun chen Barn Wien, 46-63
|
Pielou E C. 1975. Ecological diversity. New York: John Wiley and Sons, 20-45.
|
Simpson E H. 1949. Measurement of diversity. Nature: 163;688. |
Shannon C E, Weaver W. 1949. The Mathematical Theory of Communication. Urbana: University of Illinois Press.
|