 2006, Vol. 42
2006, Vol. 42文章信息
- 程诗明, 顾万春.
- Cheng Shiming, Gu Wanchun.
- 苦楝表型性状梯度变异的研究
- Studies on Phenotypical Characteristics Gradient Variation of Melia azedarach
- 林业科学, 2006, 42(5): 29-35.
- Scientia Silvae Sinicae, 2006, 42(5): 29-35.
-
文章历史
- 收稿日期:2004-10-11
-
作者相关文章



2. 中国林业科学研究院林业研究所 北京 100091
形态结构进化的总趋势是复杂性的提高和多样性的增长(沈银柱,2002)。从形态学或表型性状来检测植物遗传变异是最简便而直接的方法(葛颂等,1994)。表型多样性研究,通常集中在变异稳定的繁殖器官性状、形态性状、生理生化性状及经济生产率性状等方面(顾万春,2004)。对于大多数植物来说,花部等生殖器官性状相对于营养器官来说是比较保守和稳定的(Stebbins, 1950; Davis et al., 1963),因此,生殖器官性状受环境控制较小,是研究性状梯度变异、表型多样性的好材料。
林木种群分布多数呈随机方式沿复合环境梯度而散布, 群体间相互关系依从于环境梯度而排列(惠特克,1986;斯特恩等,1984)。每个树种以自己的遗传、生理以及对环境反应的方式而分布着。广域分布种不同群体在表型上存在很大的变异,尤其表现在物候、繁殖器官等性状上(Stebbins, 1950)。连续分布的种群,伴随着生态渐变,适应与保存了梯度变异(顾万春,1995)。表型性状的连续变异称为表型梯度(phenotypic gradient)变异。
苦楝(Melia azedarach)分布连续广泛,自南向北横跨热带、亚热带、温带,分布区地形、地貌等地理生态条件复杂,经过长期自然选择,蕴含着丰富的种内变异,形成了许多具有适合不同气候的地理生态型,具广泛的遗传基础。根据群体/家系样本分析形态性状在群体间、群体内变异大小,了解苦楝表型性状的遗传基础及其变异模式,为苦楝种质资源的保存、构建核心种质提供科学依据。
1 材料与方法 1.1 材料在全面收集苦楝分布资料和实地勘察、将我国苦楝全分布区划分为11个物候区(程诗明等,2005)的基础上,按中国生态梯度图(顾万春,1992;1993;1995)和苦楝物候区划,在保证每个物候区至少2个群体的原则下进行随机分组取样,最终在苦楝自然分布区的11个物候区内选择了24个群体729个家系。群体内家系的选择:在询问当地居民苦楝来源的情况下,尽量选择当地土生土长,人为干扰较小、生长正常、无病虫害的成年母树,母树间距保证500 m以上。于2003年11月—2004年1月分别采集苦楝各群体/家系的种子(树冠中部偏南的位置采集成熟的单果400~500个)、叶片(从枝条顶端向下数第三回羽状复叶并采集不少于60片,压制成标本)。采样群体基本覆盖了苦楝自然分布区。苦楝采样群体的地理气候条件见表 1,生长状况见表 2。
|
|

|
|

所研究的性状为叶片及单果、核果、种子等表型可塑性(phenotypic plasticity)小、主要受遗传控制的13个形态性状:叶片长、叶片宽、叶柄长、单果长、单果宽、果柄长、核果长、核果宽、种子长、种子最宽、种子较宽(垂直于种子最宽方向量取)、单果种子数、千粒质量。将试材带回实验室后立即对上述表型性状进行测量。叶片用直尺测定(精度为0.1 cm);千粒质量用电子天平测定(精度为0.01 g);单果种子数是计算每个核果出籽粒数,计数单位为个;其他各性状用游标卡尺测定(精度为0.1 mm)。共观测了24个群体729个家系,每群体每个家系的每个观测性状随机测定60次(不重复抽样),每性状共计测定729×60=43 740次。
1.2.2 分析方法1) 典范相关分析(Canonical Correlation Analysis, CCA)典型相关分析在林业上已有较多应用(Falkenhagen et al.,1978;陈晓阳,1990;陈晓阳等,1990; 顾万春,1995;杨传平等,2001)。为了了解苦楝群体表型性状梯度变异特点,探讨其变异模式,本文以各采种点地理气候因子(经度、纬度、海拔、年均温、年降水)及与其对应的苦楝18个表型性状因子:叶片长(leaf length, Ll)、叶片宽(leaf width, Wl)、叶片长宽比(leaf length/leaf width, Ll/Wl)、叶柄长(leaf bar length, Llb)、单果长(fruit length, Lf)、单果宽(fruit width, Wf)、单果长宽比(fruit length/fruit width, Lf/Wf)、果柄长(fruit bar length, Lfb)、核果长(core length, Lc)、核果宽(core width, Wc)、核果长宽比(core length/core width, Lc/Wc)、种子长(seed length, Ls)、种子最宽(seed width 1, Ws1)、种子长最宽比(seed length/seed width 1, Ls/Ws1)、种子较宽(seed width 2, Ws2)、种子长较宽比(seed length/seed width 2, Ls/Ws2)、单果种子数(seed number, Ns)、千粒质量(mass of 1 000 seeds, M), 进行典范相关分析(CCA),研究各性状地理生态变异基本模式,揭示地理生态因子与苦楝表型性状的相关关系,将多元变量降为二元坐标。
2) 主成分分析(Principal Component Analysis, PCA)在典范相关分析基础上进行主成分分析(PCA),把二元坐标降为一元数值,将苦楝分布区的表型性状梯度变异转换成1维数据轴,即表型性状梯度轴(Phenotypic Character Gradient Axes, PCGA)。计算出苦楝表型性状梯度值(Phenotypic Character Gradient Values,PCGV),采用内插法在苦楝自然分布区内按0.2表型性状梯度差值绘出等值线图。所有绘图操作都在计算机上完成。
3) 通径分析(Path Analysis)为进一步了解苦楝各表型性状对表型性状梯度值(PCGV)的直接作用和间接作用大小,对18个表型性状与其对应性状梯度值做了通径分析。所用统计分析软件:SAS7.0, SPSS11. 0, AMOS5.0, SPQG3.0,NTSYSpc2.11;做图软件:Erdas8.7,PCI9.0,Surfur8.0,Photoshop7.0等。
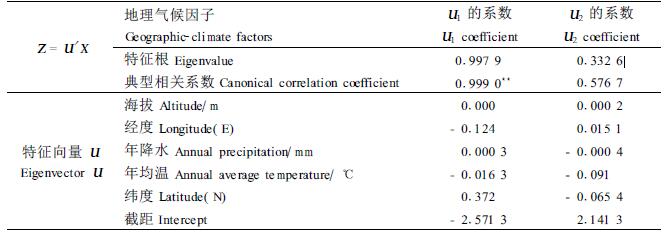
2 结果与分析 2.1 地理气候因子与表型性状的典型相关分析对苦楝群体的地理气候因子及其对应的表型性状进行典型相关分析,各因子的系数(载荷)见表 3、表 4。
|
|
|
|

从表 3中可以看出,第1对典型变量的相关系数为0.999 0,极显著(0.01水平),而表 4中第2对典型变量相关系数不显著,因此可以认为综合地理生态因子第1典型变量对苦楝群体第1综合表型性状(W1)的影响很大,而第1综合地理生态因子中起主要作用的是纬度,其次是经度。纬度的载荷为0.372,经度的载荷为0.124,因此可以认为苦楝群体表型性状受经度、纬度的双重控制,纬向为主、经向为辅的地理变异模式。叶片宽、单果长、单果宽、单果长宽比、核果长、核果宽、种子长、种子最宽、种子较宽、种子长较宽比等性状在第1典型变量中载荷最高,绝对值在1.5以上,具明显的梯度变异特点;核果长宽比、叶柄长、叶片长、种子长最宽比等性状在第1典型变量中的载荷较高,在0.8以上,也具有明显的地理变异趋势;单果种子数、千粒质量、果柄长、叶片长宽比等性状受纬度、经度控制较弱。从第1典型变量各性状的载荷符号来看,单果长、核果长、单果种子数、核果长宽比、叶柄长、叶片长、叶片长宽比、种子长、种子长最宽比、种子最宽与纬度符号相同,为正相关,表明这些性状有随纬度增高而增大的变异趋势;单果宽、单果长宽比、果柄长、核果宽、千粒质量、叶片宽、种子长较宽比、种子较宽等性状符号与经度的相一致,为正相关。因此可以得出如下结论:苦楝群体表型性状在长度上由南到北逐渐增大;在宽度上由东向西逐渐增大;南部、东部群体较北部、西部群体生长快,种粒小。
2.2 性状梯度值的计算 2.2.1 各群体第一典范变量的典范坐标第1典范变量的特征根为0.997 9,占全部特征根的78%,第1对典型变量的相关系数为0.999 0,极显著。因此,采用第1典型变量二元坐标值跟进主成分分析能代表绝大部分信息。各群体表型性状的典型坐标与性状梯度值见表 5。
|
|
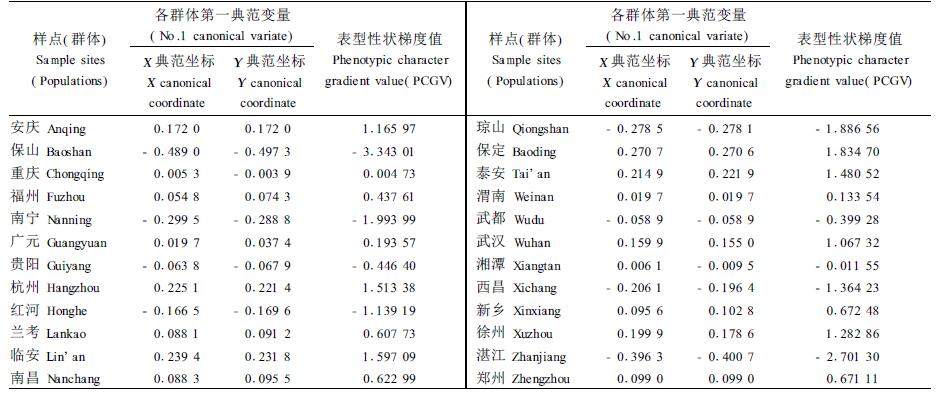
对相关分析结果中第1典型变量的二元坐标数值跟进主成分分析。第1典型变量主成分分析结果得出:第1主成分的数值变幅为-3.34~1.834 7,而第2主成分的取值范围为-0.08~0.08,第2主成分数值很小且变动幅度不大,因此,取第1主成分能代表绝大多数信息。从表 6可知,第1主成分特征根为1.999 0,贡献率高达99.95%。因此,在典型相关分析的二元变量基础上,选用第1主成分做表型性状梯度值,信息量仅损失0.05%。
|
|

利用图形图像软件,将降成一维的样本数据采用内插法将各样点连接成等值线,绘制出苦楝中国分布区表型性状等值线图(图 1),即苦楝表型性状梯度变异图。从图 1中可以很清楚地看出苦楝表型性状梯度变异的趋势特点。苦楝表型性状随着纬度、经度梯度变异明显,具很强的东北、西南变异趋势。图中海南岛的苦楝表型梯度值随纬度的减小而增大,可能与海南岛独特的地理位置分不开,该处常年受海洋气候的影响,雨量充沛、日照时数长、温度变化较缓和。从图中等值线的疏密可以看出苦楝表型多样性丰富区在云南西部、川西南和广西、广东一带,这与当地地形地貌复杂多样、气候因子变化剧烈分不开的。

|
图 1 苦楝表型性状梯度变异图 Fig. 1 The phenotypical gradient variation map of M. azedarach |
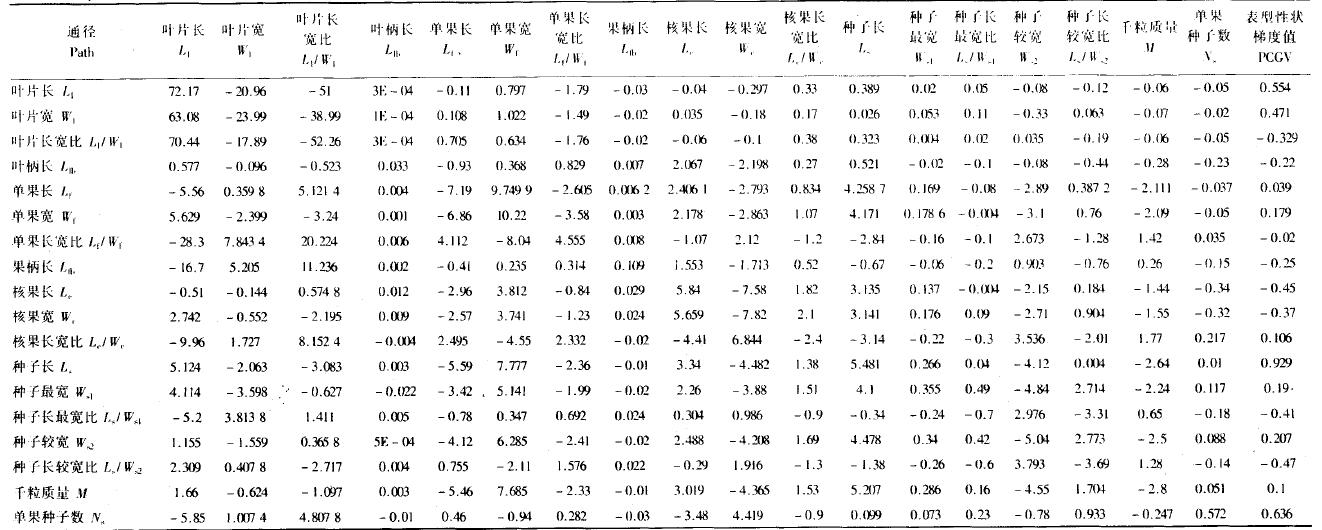
从苦楝表型性状-性状梯度值通径分析表(表 7)可以清楚地看出这些性状是如何通过直接与间接作用来影响表型性状梯度值(PCGV)的。叶片长对表型性状梯度值的直接影响高达72.17,通过叶片宽(-20.95)、叶片长宽比(-51)等其他17个性状的间接影响,加起来总的影响为0.554,即叶片长与PCGV的相关系数(其中正号表示性状之间的促进作用,负号表示抑制作用);核果长对表型性状梯度值总的作用为-0.45,起抑制作用。种子长对表型性状梯度值的直接影响为5.481,综合影响为0.929,为最大;叶柄长对表型性状梯度值的直接影响最小,为-0.033。因此,对表型性状梯度值直接影响最大的是叶片等性状,而单果、种子等繁殖器官性状普遍较小,这也从侧面说明繁殖器官性状受环境控制较小、表型可塑性小、比较稳定。
|
|
苦楝分布大半个中国,广域范围内生态梯度明显。本文以典型相关分析跟进主成分分析将多元表型性状降成1维数据轴,勾绘出苦楝表型性状梯度变异等值线图,直观明了地展示了苦楝表型性状梯度变异的地理模式。
苦楝表型性状具很强的东北、西南方向趋势变异特点:在长度性状上由南到北逐渐增大;在宽度性状上由东向西逐渐增大;南部、东部群体较北部、西部群体生长快,种粒小。
从苦楝表型性状等值线图的疏密可以清楚地展现苦楝表型多样性富集区为西南云贵川与华南广西、广东一带。为苦楝遗传多样性保存提出优先保存区域。
用通径分析的方法揭示了苦楝各表型性状对表型性状梯度值直接、间接的影响,并从侧面阐明了繁殖器官性状相对稳定性的事实。
程诗明, 顾万春. 2005. 苦楝中国分布区的物候区划. 林业科学, 41(3): 186-191. DOI:10.3321/j.issn:1001-7488.2005.03.032 |
陈晓阳. 1990. 典型相关分析在树种地理变异研究中的应用. 北京林业大学学报, 12(3): 53-60. |
陈晓阳, 陈振丙, 吴栓柱, 等. 1990. 侧柏种苗性状地理变异及气候生态基础. 河南林业科技, (1): 5-10. |
顾万春. 2004. 统计遗传学. 北京: 科学技术出版社.
|
顾万春. 1995. 中国林木育种区. 北京: 中国林业出版社, 35-127.
|
顾万春. 1993. 生态梯度轴(EGA)用于林木生态遗传的研究—CA方法估算EGA(CA1)和EGA(r2). 生态学报, 13(2): 148-156. DOI:10.3321/j.issn:1000-0933.1993.02.007 |
顾万春. 1992. 生态梯度轴(EGA)用于林木生态遗传的研究—PCA方法估算EGA(PCr). 生态学报, 12(4): 332-340. |
葛颂, 洪德元. 1994.遗传多样性及其检测方法//中国科学院生物多样性委员会.生物多样性研究的原理与方法.北京: 中国科学技术出版社, 123-140
|
惠特克R H.1986.植物群落排序.王伯荪, 译.北京: 科学出版社, 52-1142
|
沈银柱. 2002. 进化生物学. 北京: 高等教育出版社.
|
斯特恩K, 罗奇L.1984.森林生态系统遗传学.毛士田, 等译.北京: 中国林业出版社, 296-311
|
杨传平, 刘桂丰. 2001. 长白落叶松种群地理变异研究. 应用生态学报, 12(6): 801-805. DOI:10.3321/j.issn:1001-9332.2001.06.001 |
Falkenhagen E R, Nash Sr W. 1978. Multivate classfication in provenance research. Silvae Genetic, 21(1): 14-23. |
Stebbins G L. 1950. Variation and evolution in plants. New York: Columbia Univ Press.
|
Davis P H, Heywood V H. 1963. Principles of angiosperm taxonomy. Edinburgh and London: Oliver & Boyd.
|