 2006, Vol. 42
2006, Vol. 42文章信息
- 孙守家, 古润泽, 丛日晨, 车少臣, 高俊平.
- Sun Shoujia, Gu Runze, Cong Richen, Che Shaochen, Gao Junping.
- 银杏树干茎流变化及其对抑制蒸腾措施的响应
- Change of Trunk Sap Flow of Ginkgo biloba and Its Response to Inhibiting Transpiration
- 林业科学, 2006, 42(5): 22-28.
- Scientia Silvae Sinicae, 2006, 42(5): 22-28.
-
文章历史
- 收稿日期:2004-12-23
-
作者相关文章



2. 北京市园林科学研究所 北京 100102
2. Beijing Institute of Landscape and Garden Beijing 100102
随着我国城市建设速度的不断加快,城市绿化特别是道路绿化已打破春秋植树的惯例,随时和不定期的移植日益普遍。移植过程中树体水分失衡是制约树木移植成活的一个关键因素(Hipps et al., 1996;1999)。因此,对水分运输规律和水分调控技术的研究已成为一个迫切的问题。通过精确地测定茎流速率,能够确定植物水分运输的规律(Fernández et al., 2001),对于验证抑蒸节水措施具有重要意义。利用热脉冲方法,先后对杨树(Populus deltoides)(刘奉觉等,1993)、棘皮桦(Betula dahurica)和五角枫(Acer mono)(李海涛等,1998)、小美旱杨(Populus popular)(高岩等,2001)、胡杨(Populus euphratica)和柽柳(Tamarix spp.)(张小由等,2003)、胡杨幼树(司建华等,2004)的树干茎流进行研究,但是银杏树干的茎流变化规律尚未见报道。
本研究以在城市绿地和道路绿化中广泛应用的银杏(Ginkgo biloba)为试材,通过监测树干茎流时空动态变化来分析银杏水分运输的规律,并研究了摘叶、遮荫、喷抗蒸腾剂等抑制蒸腾措施对其水分运转的影响,为进一步开展大规格苗木移植过程中树体水分控制技术研究提供理论依据。
1 材料与方法 1.1 试验地概况试验在北京市园林科学研究所内进行。试验地所在地区属于温带半湿润大陆性季风气候,年平均气温11~12 ℃,年极端高温在38~40 ℃,极端低温在-14~-20 ℃,年降雨量在500~600 mm,且集中在6—9月,年均风速2.7 m·s-1,年蒸发量1 956.8 mm,年平均总日照时数2 554.8 h,无霜期190~195 d。
1.2 试验材料银杏15年生,长势良好,树干通直无明显疤痕,树盘铺有5 cm厚的松树皮,树下种植早熟禾草坪,每周喷灌浇水2次,生长环境良好。选择干形通直、冠形良好的银杏树4株,具体参数详见表 1。在银杏树干上位(树冠基部,距地表面2.2 m)、中位(胸径,距地表面1.3 m)和下位(树干基部,距地表面0.2 m)3个位点测定不同高度茎流变化,深度为形成层以下10 mm。不同深度的茎流测定是在树干正南方,深度分别为形成层以下5、10、15、20 mm,高度为1.3 m;不同方位的茎流测定分别在树的东、南、西、北方,高度为1.3 m,深度为形成层以下10 mm。
|
|
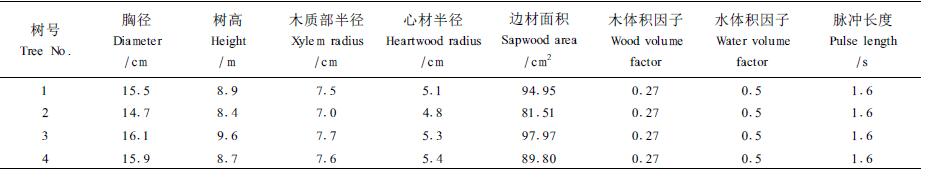
茎流测定用澳大利亚联邦科学与工业研究组织(CSIRO)开发的GREENSPAN茎流测定系统(Sap Flow System)。数据采集器(data-logger)连接4个SF-300分体式探头,每个探头具有1个热探针和2个感应探头,热探针长90 mm,感应探头长80 mm,直径2 mm。将随机提供的模具紧贴树干,用2 mm钻头的电钻水平钻孔。探头用铝箔包裹防止太阳辐射,胶带密封防止雨水进入。数据采集器与探头连接,接通12 V电源,与建立超级终端的电脑笔记本相连,设置数据采集参数,采集间隔期为20 min,脉冲时长为1.6 s;同时设置木质部半径、心材半径、木体积因子、水体积因子等参数。使用随机提供的SAPPRO分割数据文件,SAPCAL软件进行数据处理,同时计算茎流速率、茎流通量和累积茎流通量。
气象因子用美国Onset Computer Corporation公司的HOBO气象站测定,设置数据采集2 min读数1次,20 min将所读数据平均并记录,同步监测树冠中部(距地表面4 m)空气温度、空气相对湿度、太阳辐射、有效辐射、风向、风速,以及地下20 cm的土壤温度和土壤湿度,使用随机提供的BOXCAR Pro4软件进行数据处理。
遮荫处理使用单层黑色遮荫网遮荫,用ST-85自动量程照度计测定透光率为46.2%;北京市园林科研所生产的抗蒸腾剂,稀释20倍整株叶面喷施;剪叶处理大约摘掉整个树冠的2/3叶子。
测定时间2004年8—9月。数据使用MICROSOFT EXCEL和SAS 8.0分析软件进行处理。
2 结果与分析 2.1 银杏树干茎流在不同部位之间的变化 2.1.1 树干茎流的轴向变化从图 1可看出树干不同高度的木质部具有不同的茎流速率,上位茎流速率大于下位茎流速率,并且白天茎流速率差异更显著。造成差异的原因可能是树干上部直径小,边材面积小,在相同水分运输通量的情况下,上部边材茎流速率快,下部边材茎流速率慢。从树木解剖学特征看,木质部导管的直径自下而上逐渐减小,随着测定的位置升高,被测部位的导管直径越来越小,在通过相同水量的情况下,茎流速率也随之增大。通过树体上位、中位和下位截面的累积茎流通量差异不明显(图 1),这说明树干不是耗水的主要部位,存在这种差异可能与树木本身的水力结构和水容性有关(Goldstein et al., 1998)。

|
图 1 银杏树干上位、中位和下位的茎流速率、累积茎流通量变化动态 Fig. 1 Changes of sap flow velocity and cumulative flux at different positions of G. biloba trunk |
从图 2可以看出4个观测位点的茎流速率变化趋势相近,但是在径向深度上存在明显的差异。10 mm位点茎流速率最大,5 mm位点和15 mm位点基本相同,20 mm位点最小。其主要原因可能是木质部边材在结构上的异质性导致不同深度的木质部在导水能力上存在差异(Zimmermann,1983)。

|
图 2 银杏树干不同深度茎流速率变化 Fig. 2 Changes of sap flow velocity at different depths of G. biloba trunk |
从图 3可以看出,树干东南西北4个位点的茎流速率具有相同的变化趋势,但是其变化幅度差异很大。其中,南侧的茎流速率最大,特别是在辐射较强的晴天差异更加明显;东侧和西侧的茎流速率差别不明显;北侧的茎流速率最小。

|
图 3 银杏树干不同方位茎流速率变化 Fig. 3 Changes of sap flow velocity at different directions of G. biloba trunk |
从图 4可看出,晴天(8月1日)茎流速率呈现准单峰曲线变化,夜晚有较小而稳定的茎流速率,凌晨6:00开始启动急速增高,10:00后增速缓慢并逐渐稳定,15:30开始又迅速降低,19:30后逐渐稳定在较低的水平上,茎流属于骤升骤降型;多云天气(8月10日)条件下,由于太阳辐射等气象因子的波动造成茎流速率变化不稳定,呈多峰曲线变化,且白天茎流速率要远小于晴天;阴雨天气(8月12日)条件下,由于白天降雨,茎流速率与夜间相比变化不大。由此可见,不同天气条件下,银杏树干茎流存在显著差异。

|
图 4 银杏树干在不同天气状况下的茎流速率日变化 Fig. 4 Daily changes of sap flow velocity of G. biloba trunk under different weather conditions |
图 5是3种不同天气(8月1日、3—6日晴朗, 8月2日阴雨,8月7—8日多云)情况下银杏茎流速率的日际变化特征,可以看出白天茎流速率大于晚上。晴天茎流速率比其他天气大,且早晨启动较早晚上停滞较晚,大约在凌晨6:00启动19:30停滞;阴雨和多云天气茎流速率较小且启动较晚停滞较早,大约在6:30左右启动16:00停滞。8月2日中午下雨以后空气湿度较大,与晴朗天气相比白天茎流速率小。晚上树体依然存在较小的茎流,这并不是晚上植物蒸腾耗水,而是晚上根压存在推动树液流动,补充白天植物蒸腾耗掉的水分。

|
图 5 银杏树干茎流速率的日际变化(2004年8月1—8日) Fig. 5 Changes of sap flow velocity of G. biloba trunk from 1st to 8th August, 2004 |
从图 6可以看出,晴天树干茎流与太阳辐射变化趋势大体相同,早晨茎流启动的时间和太阳辐射启动的时间基本一致, 主要是太阳辐射控制气孔的活动。多云及阴雨天气较低的太阳辐射对应多云及阴雨天气较低的茎流速率。

|
图 6 银杏茎流速率与有效辐射、气温、相对湿度、风速、降雨量等环境因子之间的关系(2004年8月4—11日) Fig. 6 Correlation between sap flow velocity and environmental factors including photosynthetically active radiation, temperature, relative humidity, wind speed, and rainfall during 4th to 11th, August 2004 |
晴天气温在凌晨5:00左右达到最低点,随后开始启动,时间大约比树干茎流启动时间晚0.5 h左右,15:00左右达到最高点。相对湿度和树干茎流变化的关系与气温和树干茎流变化的关系恰好相反。气温的波峰和相对湿度波谷与树干茎流的午后高峰基本重合,这可能是此时太阳辐射下降,气孔重新开放,在温度和湿度的共同影响下使得叶内外的蒸汽压差加大,有利于水分从叶内逸出,蒸腾作用加强,导致树干茎流速率升高。茎流速率在太阳有效辐射下降时而没有立刻降低,这种差异可能是由于气温和相对湿度与太阳辐射的不同步性造成的。
从图 6中可以看出白天的风速较大,夜晚的风速较小,与树干茎流变化有一定的关系,但是风向对树干茎流的影响有待进一步研究。
阴雨天茎流速率保持低值。8月9、11日有2次降水过程。在降雨时茎流速率迅速下降,降雨结束后快速上升,形成一个较大波谷。主要是降雨增加了空气相对湿度,极大地降低了叶片气孔内外的蒸汽压梯度。
2.3.2 逐步回归分析建立树干茎流估算模型通过以上分析,对于3种不同天气(8月2—16日)的树干茎流速率(y)以及影响其变化的环境因子土壤温度(x1)、降雨量(x2)、风速(x3)、风向(x4)、土壤含水量(x5)、大气压(x6)、气温(x7)、空气相对湿度(x8)、太阳有效辐射(x9)等进行逐步回归分析,详见表 2。
|
|
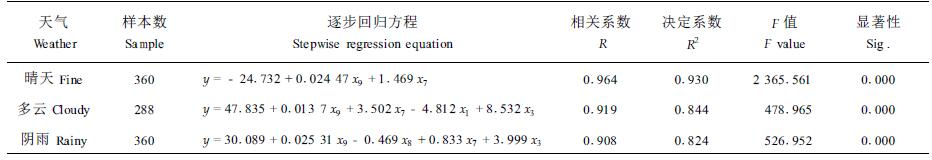
从表 2中可以看出,3个方程的相关系数R都大于0.9,决定系数R2都在0.8以上,方程都达到了极显著水平。3种天气情况下太阳有效辐射x9、气温x7都能显著地影响树干茎流变化。多云和阴雨天气风速也是主要的影响因子之一,它能够将气孔外边的水蒸气吹走,减小叶片边界层阻力,影响茎流的变化。阴雨天气中由于降水使得空气相对湿度变大,因此相对湿度x8也是影响茎流速率的一个重要因素。
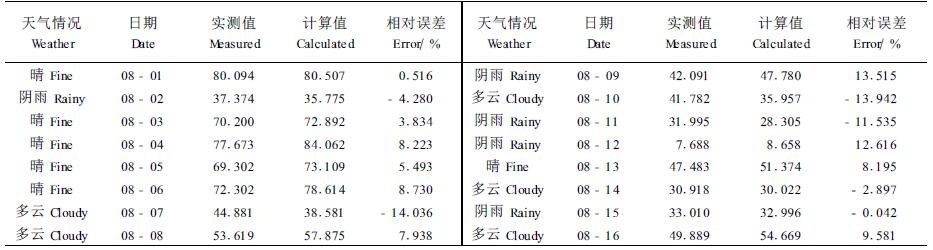
利用上述逐步回归方程对8月1-16日中午12:00茎流速率进行了验证,结果详见表 3。从中看出,由回归方程求算的理论茎流速率值与实际测得的结果之间基本吻合,最大的相对误差是14.04 %,最小相对误差是0.04 %。这说明使用较为简洁的逐步回归方程来预测茎流的变化是可行的。
|
|
从图 7可以看出,第1天茎流速率相差较小,从第2天10:00开始进行不同处理,遮荫处理后白天的茎流速率明显下降,大约只有前一天的1/3;剪叶处理后白天的茎流速率下降比较明显,比前一天大约下降了1/2;喷抗蒸腾剂处理后白天的茎流速率略有下降,大约下降了10%。晚上各处理和对照之间的茎流速率变化差别不明显,主要是因为晚上蒸腾耗水基本停止,只有根压维持树体水分流动。

|
图 7 不同蒸腾抑制措施对银杏树干茎流速率的影响(2004年8月28—31日) Fig. 7 Effect of different treatments of inhibiting transpiration on sap flow velocity of G. biloba trunk during 28th to 31st August 2004 |
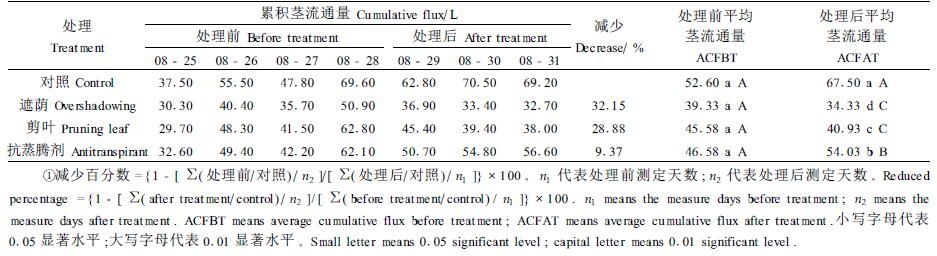
从表 4每天的累积茎流通量变化可以看出,8月29日上午10:00开始处理后,遮荫、剪叶和喷抗蒸腾剂处理后的累积茎流通量和对照相比明显下降,其中遮荫和剪叶处理效果最明显,分别下降32.15%、28.88%,喷抗蒸腾剂处理后累积茎流通量下降9.37%。对4棵树木样本的累积茎流通量进行单因素方差分析并用最小显著差数法(LSD)检验,从表 4可以看出,处理前各棵树的累积茎流通量无论在0.05水平还是在0.01水平上差异都不显著,说明试验材料比较一致;处理后各棵树之间的累积茎流通量在0.05水平上差异显著,在0.01水平上抗蒸腾剂、遮荫和剪叶处理与对照差异极显著,剪叶和遮荫处理之间差异不显著。这说明采取遮荫、剪叶、喷抗蒸腾剂处理都可以有效地抑制叶片蒸腾耗水,遮荫和剪叶处理的效果比喷抗蒸腾剂更明显。
|
|
水分的运输主要在边材的木质部进行。Edwards和Booker(1984)采用甲苯胺蓝染色法对杨树树干的径向水力导度进行研究表明,在木质部边材区不同深度的位点之间的透水性差别较大。Edwards等(1996)提出二次曲线理论,认为在木质部茎流随木质部深度的变化遵从二次曲线的关系。银杏木质部茎流速率并不随着木质部深度增加逐渐减小,而是在10 mm位点茎流速率最大,5 mm和15 mm差别较小;这与对樟子松(Pinus sylvestris var. mongolica)观测的结果(吴丽萍等,2003)一致,而与孙鹏森等(2000)在油松(Pinus tabulaeformis)上观测的结果不同。存在这种差异的原因可能是不同树种的边材在结构上存在差异,从而导致木质部在导水能力上存在差别(Zimmermann,1983)。不同方位木质部茎流速率差异显著,南侧茎流速率最大,北侧茎流速率最小。造成这种现象的原因可能有2个:一是不同方向的木质部生长的速度不同,位于南侧的木质部生长较快,导管系统发达通畅;二是光线的直射点落在南侧的时间较长。这也表明树干茎流运动是沿着导管直线上升的,而非螺旋上升(龚道枝等,2001)。上位、中位和下位的累积茎流通量差异不显著,这与王华田等(2002)在油松上观测的结果差别较大,主要是因为本文所测的树干上位是在树冠基部,这段区域没有分枝和叶片。这也证明树干不是耗水的主要部位,大部分水被叶片蒸腾消耗。
水分在植物体内的运输是一个复杂的过程,既由自身生理特性决定(Gullo et al., 1992),同时也与太阳有效辐射、气温、空气相对湿度、风速、土壤含水量、土壤温度、大气压等环境因子密切相关(丁日升等,2004),这些因子相互作用、相互协调,共同影响树干茎流变化。在典型的晴天、多云、阴雨天气下,银杏树干茎流速率的日变化差异显著。晴天茎流启动较早,峰值大;多云天气茎流白天呈现不规则的多峰变化,规律性不强;阴雨天气茎流保持低值,启动晚,达到峰值后逐渐下降。茎流也存在显著的日际变化,白天茎流速率大于晚上。这主要是因为白天叶片的蒸腾量较大,蒸腾拉力是茎流传输的主要驱动力;晚上茎流虽然很小但依然存在,主要是根压作用(Clark et al., 1957)推动树液流动,补充白天植物蒸腾耗掉的水分,恢复植物体内的水分平衡。逐步回归分析表明3种天气(晴天、多云、阴雨)情况下太阳有效辐射、气温都是影响树干茎流变化的主导因子。多云和阴雨天气风速也是主要的影响因子之一,它能够将气孔外边的水蒸气吹走,减小叶片边界层阻力,影响叶片的蒸腾。阴雨天气中相对湿度也是影响茎流速率的一个重要因素。
树木移植过程中树体水分失衡导致的干旱胁迫,是制约树木移植成活的一个重要因素,切断根系降低了吸收水分的能力(Hipps et al., 1996;1999),尽可能地减少水分散失是保证树木成活的关键。差异显著性检验表明遮荫、剪叶和喷抗蒸腾剂等措施可以有效地抑制叶片蒸腾耗水。遮荫和剪叶处理效果比较明显,能够降低30%左右的蒸腾耗水,但在应用中比较耗工耗时;喷抗蒸腾剂大约能降低10%的蒸腾耗水,应用比较方便。因此,在树木移植尤其是在生长季节的移植过程中,3种抑制蒸腾措施综合使用,能够缓解水分失衡导致的胁迫程度,提高生长季节树木移植的成活率。
丁日升, 康绍忠, 龚道枝. 2004. 苹果树液流变化规律研究. 灌溉排水学报, 23(2): 21-25. |
高岩, 张汝民, 刘静. 2001. 应用热脉冲技术对小美旱杨树干液流的研究. 西北植物学报, 21(4): 644-649. DOI:10.3321/j.issn:1000-4025.2001.04.007 |
龚道枝, 王金平, 康绍忠, 等. 2001. 不同水分状况下桃树根茎液流变化规律研究. 农业工程学报, 17(4): 34-38. DOI:10.3321/j.issn:1002-6819.2001.04.008 |
李海涛, 陈灵芝. 1998. 应用热脉冲技术对棘皮桦和五角枫树干液流的研究. 北京林业大学学报, 20(1): 1-6. DOI:10.3321/j.issn:1000-1522.1998.01.001 |
刘奉觉, Edwards W R N. 1993. 杨树树干液流时空动态研究. 林业科学研究, 6(4): 368-372. DOI:10.3321/j.issn:1001-1498.1993.04.018 |
司建华, 冯起, 张小由. 2004. 热脉冲技术在确定胡杨幼树干液流中的应用. 冰川冻土, 26(4): 503-508. DOI:10.3969/j.issn.1000-0240.2004.04.019 |
孙鹏森, 马履一, 王小平, 等. 2000. 油松树干液流的时空变异性研究. 北京林业大学学报, 22(5): 1-6. DOI:10.3321/j.issn:1000-1522.2000.05.001 |
王华田, 马履一, 孙鹏森. 2002. 油松、侧柏深秋边材木质部液流变化规律的研究. 林业科学, 38(5): 31-37. DOI:10.3321/j.issn:1001-7488.2002.05.006 |
吴丽萍, 王学东, 尉全恩, 等. 2003. 樟子松树干液流的时空变异性研究. 水土保持研究, 10(4): 66-68. DOI:10.3969/j.issn.1005-3409.2003.04.018 |
张小由, 龚家栋, 周茂先, 等. 2003. 应用热脉冲技术对胡杨和柽柳树干液流的研究. 冰川冻土, 25(5): 585-589. DOI:10.3969/j.issn.1000-0240.2003.05.017 |
Clark J, Gibbs R D. 1957. Studies on tree physiology Ⅳ: Further investigation of seasonal changes in moisture content of certain Canada forest tree. Canadian Journal of Botany, 35: 219-253. DOI:10.1139/b57-021 |
Edwards W R N, Becker P. 1996. A unified nomenclature for sap flow measurements. Tree Physiology, 17: 65-67. |
Edwards W R N, Booker R E. 1984. Radial variation in the axial conductivity of Populus and its significance in heat pulse velocity measurement. Journal of Experimental Botany, 33(153): 551-561. |
Fernández J E, Palomo M J, Díaz-Espejo A, et al. 2001. Heat-pulse measurements of sap flow in olives for automating irrigation: tests, root flow and diagnostics of water stress. Agriculture Water Management, 51: 99-123. DOI:10.1016/S0378-3774(01)00119-6 |
Goldstein G, Andrade J L, Meinzer F C, et al. 1998. Stem water storage and diurnal patterns of water use in tropical forest canopy trees. Plant Cell and Environment, 21: 397-406. DOI:10.1046/j.1365-3040.1998.00273.x |
Gullo M A, Salleo S, Lo Gullo A. 1992. Water storage in the wood and xylem cavitation in 1-year-old twigs of Populus deltoids Bartr. Plant, Cell and Environment, 15: 431-438. DOI:10.1111/j.1365-3040.1992.tb00993.x |
Hipps N A, Higgs K H, Collard L G. 1996. The effect of irrigation and root pruning on the growth of sycamore (Acer pseudoplatanus) seeding in nursery beds and after transplantation. Journal of Horticulture Science, 71: 819-828. DOI:10.1080/14620316.1996.11515464 |
Hipps N A, Higgs K H, Collard L G. 1999. Effects of root wrenching on the growth and water relations of Prunus avium and Castanea sativa seeding in nursery beds and after outplanting. Canadian Journal of Forest Research, 29(6): 696-704. DOI:10.1139/x99-044 |
Zimmermann M H. 1983. Xylem structure and the ascent of sap. Berlin: Springer Verlag.
|