 2006, Vol. 42
2006, Vol. 42文章信息
- 李伟成, 盛海燕, 潘伯荣, 常杰.
- Li Weicheng, Sheng Haiyan, Pan Borong, Chang Jie.
- 3种沙漠植物地上部分形结构与生物量的自相似性
- Self-Similarity Relationship between Component of Shoot and Biomass of Three Hungriness Plants
- 林业科学, 2006, 42(5): 11-16.
- Scientia Silvae Sinicae, 2006, 42(5): 11-16.
-
文章历史
- 收稿日期:2004-08-05
-
作者相关文章


2. 杭州环境保护科学研究院 杭州 310005;
3. 中国科学院新疆生态与地理研究所 乌鲁木齐 830011;
4. 浙江大学生命科学学院 杭州 310029
2. Hangzhou Environmental Protection Science Institute Hangzhou 310005;
3. Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences Ulmuqi 830011;
4. College of Life Science, Zhejiang University Hangzhou 310029
目前,荒漠化与盐碱化是人类面临的2个环境问题,在塔里木盆地的塔克拉玛干沙漠表现得尤其突出(陶玲等,2001;李伟成等,2003)。塔克拉玛干地区生态环境恶劣,气候极度干旱,因此一些抗热、抗寒、抗强碱的优良固沙先锋植物在这里就显得尤为重要了(潘伯荣等,1994;陶玲等,2001)。柽柳(Tamarix spp.)作为干旱、半干旱区广泛分布的灌木,具有强大的固定流沙和耐盐碱能力;梭梭(Haloxylon ammodendron)是我国干旱荒漠区的优良固沙植物,是荒漠濒危类3级国家保护植物(陶玲等,2001);而沙拐枣(Calligonum mongolicum)以其独特的生活型、表型特征和薪柴、饲用、固沙、蜜源等经济特点,早就引起了人们的注意。上述3种植物具备了荒漠先锋种的生理、形态特征(潘伯荣等,1994;陶玲等,2001),是西北荒漠生态系统的关键物种。
自从Burrough(1981)将Mandelbort(1979)提出的分形和分维概念应用到自然生态和环境学科领域以来,分形理论已经在植物进化模拟(Nicklas,1986)、农作物生物量与时间尺度上的自相似性(self-similarity)(Valdez-Cepeda et al., 1998)、面积和物种丰富度的分形维数等方面进行了探讨(Harte et al., 1999)。目前,普遍认同林木的生长具有自相似性(Peitgen et al., 1991;常杰等,1995;李伟成等,2003)。本文选择这3种植物作为研究对象,基于自相似理论在各分形单元上分析比较柽柳、梭梭、沙拐枣植株在不同生境中的生物量积累,希望利用分形理论来阐明不同物种在不同生境中的自相似性。
1 研究地自然概况塔克拉玛干沙漠腹地年均降水量11.05 mm,年蒸发量3 638 mm,平均气温12.4 ℃,一年中7月最热,平均气温28.2 ℃,12月最冷,平均气温-8.1 ℃,年最高气温45.6 ℃,年最低气温-22.2 ℃,≤10 ℃年积温4 530 ℃,年均相对湿度29.4%;吐鲁番盆地年均降水量16.4 mm,年蒸发量2 727~3 837 mm,年平均气温13.9 ℃,最高气温47.6 ℃,最低气温-16. 2 ℃,≤10 ℃年积温5 391.3 ℃,年均相对湿度41%。
2 材料与方法2001年6月在塔克拉玛干沙漠运输公路腹地(39°00′N、83°40′E,海拔1 099 m)采集分布广、株型完整、无病虫害的沙拐枣、柽柳和多年生梭梭植株(人工移栽)。吐鲁番地区(42°51′N、89°11′E,海拔-95~-76 m)虽亦具典型的大陆性沙漠气候特征,但与塔克拉玛干沙漠腹地相比,地下水含量较丰富,植物生境比较优越。当月于吐鲁番盆地植物园采集上述3种植株,以比较地上部生物量在不同生境的积累情况。
分别测量2个地区植株的主茎长、分枝长、冠幅和外形体积,并采下茎、枝和叶,经80 ℃烘干48 h,分别得到茎、枝和叶干质量等性状参数,通过累加分枝生物量获得地上总质量。
为了方便,对3种植物统一假设变量。设地上部生物量为Mst,茎干质量为Ms,枝干质量为Mb,叶干质量为Ml;主茎长为Hs,分枝长为Hb,冠幅为C,外形体积为V。并有以下关系:
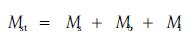
|
(1) |
在Kaye(1994)和Mandelbort(1979)的专著中均提及用分形几何的概念对树木的生物形态结构进行描述。用网格法来定量研究茎、枝、叶的分形结构时发现用分形维数可以描述各部分不同的空间占据能力,并存在如下关系:

|
(2) |
式中:M为网格覆盖区域边长(特征尺度指标);f(M)为是被覆盖网格的质量函数;D为欧氏维数;δ为分形维数。
假设S为测度指标(如质量、重量、生物量等),M为测量所采用的尺度指标(如长度、面积、体积等),δ为指数,C0、C1为常量。如存在如下关系:
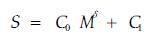
|
(3) |
则称δ为分形维数。δ可以单纯的用来表示一个系统的破碎复杂性,也可以用来描述一个整体的空间占据能力或状态(Mandlebrot, 1979;Harte et al., 1999;马克明等,2000;李伟成等,2003)。
自相似统计模型拟合结果与SPSS的逐步回归(stepwise regression)和BP神经网络模型的效果进行比较。神经网络BP算法用于多层网络,网络中不仅有输入层节点及输出层节点,而且还有一层至多层隐含层节点。本文采用具有3个输入节点(即输入节点分别是枝长、冠幅和外形体积3个参数)和单输出节点的3层BP神经网络;隐含层节点设为输入层节点数的75 %,即隐含层2个节点,输出层1个节点(即3-2-1BP神经网络模型),允许误差e<0.000 1, 迭代次数为1 000,Sigmoid参数0.9,数据进行标准化转换。
3 结果与分析 3.1 统计自相似系统植物分枝作为一个独立的分形结构,与植物地上部分是一个统计自相似系统。根据生物体生长中的分形原理,生物体自身变化应是它的分形特性在外表的显露(Nicklas,1986)。通过对塔克拉玛干腹地、吐鲁番盆地采集的柽柳、梭梭、沙拐枣植株分枝长、地上部的尺度指标(冠幅、体积)与地上部生物量做双对数处理后进行拟合(表 1),符合分形公式(3),整个地上部与分枝之间存在着良好的统计自相似性(P<0.01),表明分枝与整个地上部在空间上具有同一发展趋势,分枝是整个地上部的分形单位。同时这种统计自相似系统也给测量植物生物量提供了新的路径,分枝(子系统)与地上部(母系统)之间具有自相似性,利用其主茎或枝长的数量性状即可判断植物的地上部生物量。
|
|

在极端的物候环境下,研究先锋物种的生物量是评价先锋物种改良土壤、改善荒漠地区环境的重要指标。由表 1可以发现:塔克拉玛干沙漠与吐鲁番盆地的2年生柽柳生物量差异显著,Mann-Whitney U-test值为1.55(P<0.05);多年生梭梭和二年生沙拐枣与柽柳植物比较差异更显著,U-test值分别为29.57和35.58(P<0.01)。
在分形几何学意义下,分形维数是生物量在不同器官中累积规律(生物量分配)的表征(Waliszewski et al., 2001)。在综合指标——外形体积上,不同地区的柽柳、梭梭和沙拐枣表现出了很大的差异,在塔克拉玛干腹地,体积分形维数为梭梭<柽柳<沙拐枣,在吐鲁番盆地却是沙拐枣<梭梭<柽柳(表 1)。从生物量积累角度分析,在塔克拉玛干沙漠,沙拐枣的空间占据能力比其他2种植物强,而在吐鲁番盆地中则柽柳较强;从压缩的一维(枝长)和二维(冠幅)空间中的生物量积累来看,塔克拉玛干沙漠中梭梭在线性空间的延伸能力较强,分形维数为3.370,柽柳在压缩的冠幅空间中占据能力最强,分形维数为2.964;而吐鲁番盆地中沙拐枣和柽柳分别在一维和二维空间中表现较好,分形维数分别为2.807和2.757,表明沙拐枣以高生长为主,而柽柳以水平延展为主。
上述差异反映了植被分形结构对异质环境的生长响应(growth response)。塔克拉玛干沙漠和吐鲁番盆地柽柳、梭梭、沙拐枣3种植物的生物量存在明显的差异,表现出在逆境中植物空间占据能力较强的特性。相对塔克拉玛干沙漠而言,吐鲁番盆地能以地下水浇灌上述3种物种,风沙侵扰也较少,具备了良好的生境条件。比较两地3种植物冠幅指标的分形维数可以发现:柽柳的空间占据能力在塔克拉玛干沙漠上表现较强;梭梭在塔克拉玛干沙漠上寿命较长,所以高生长尤为突出;沙拐枣与梭梭类似,以高生长为主,在线性空间中生物量积累在吐鲁番盆地上表现较好,但是体积指标显示塔克拉玛干沙漠的沙拐枣空间占据能力较强。分形维数表明2年生柽柳、沙拐枣的枝长分形维数(δL)与冠幅分形维数(δC)相反,如2年生柽柳、沙拐枣进行高生长,则冠型缩小,反之冠幅较大则枝长较短;由于塔克拉玛干腹地的梭梭生长年限明显长于吐鲁番盆地,而(δL)和(δC)也一致表现为塔克拉玛干腹地种群的枝长及冠幅占优势。
3.3 不同模型之间的拟合结果比较分别用枝长、冠幅和外形体积3个自变量进行自相似模拟,多元逐步回归和3-2-1 BP模拟(表 2),发现3种植物用这2种方法回归模拟都达到了极相关程度。但从模拟的生物量χ2差异适合性检验分析得到:在塔克拉玛干沙漠和吐鲁番盆地中,梭梭的生物量与自相似模拟生物量的差异显著;柽柳的拟合结果是吐鲁番盆地好于塔克拉玛干沙漠(枝长、体积的χ2值分别为0.999 7和0.976 2),并且优于BP拟合(χ2=0.925 7);沙拐枣仅在吐鲁番盆地的体积χ2值达到0.373 8,比BP拟合效果要好(χ2=0.000 7)。之所以出现这种情况是因为指数形式的统计自相似模型在误差上也有缩放现象,拟合效果出现了两极分化,在样本的小数区域适合性很好,但是在大数区域呈发散状,误差较大。所以吐鲁番盆地幼龄沙拐枣和柽柳的某些参数达到了极相关,而塔克拉玛干沙漠中此两物种的检验结果为没有相关性,这是因为数据中有主导误差的相对大数出现。BP的误差逆传播减少了这种情况,能实现映射变换的前馈型的网络(Waliszewski et al., 1993),但是由于环境异质性过于强烈、人为测量的误差和研究物种的形体可塑性等因素,建模数据与检验数据差异性也很大,所以造成BP的拟合效果较差。SPSS多元回归,相关性指数R值比自相似统计模型好,但R值上升的同时,检验结果却在小数区域出现了负值,这对于生物量而言是不切合实际的,而逐步回归也表明生物量最终倾向于单一参数的线形回归。
|
|

林木生长具有自相似性,植物分枝与植物地上部分是一个统计自相似系统。但实际上,在自然界不存在严格的自相似性(Mandlebrot, 1979)。运用分形几何原理计算得出两地3种沙漠先锋种在各尺度指标上的分形维数,分形维数是对生物量在不同器官中累积规律(生物量分配)的表征,可以从线、面和空间尺度分析物种生长发育及其对生境的可塑性响应。
自相似统计模型根据不同地理、气候、环境特征回归,得到常量系数后,可以由测量的性状参数来比较地上生物量大小(李伟成等,2003),而且在一定程度上拟合检验结果准确度较高。以往在预测生物量时大多采用多元回归,其处理的数据统计差异性较小,效果较好,但是本研究发现多元回归模型在拟合此3种植物的生物量时效果比自相似模型差,并且处理差异性显著的生物量数据(有极大数和极小数)时,预测结果往往出现负值,不符合自然逻辑,说明传统的用多元回归方法进行判断和预测差异显著的沙漠植被的生物量有待商榷。3-2-1 BP非线性网络模型的拟合结果较均匀,而且结果比自相似统计模型和多元回归显著。故在模拟幼龄沙漠植被的生物量时如果数据方差较小则可以考虑运用统计自相似模型或者多元回归,而BP模型对于幼龄和多年生的数据(方差大)都比较适合。分形模型精确度不如BP网络模型,但是比静态回归模型和非线性预测模型更有信息量,可以比较物种在线性、平面和空间的占据能力,但是更多的是以一种统计指数的形式出现,与经典的分形维数在尺度上的稳定性优势有本质的区别(邬建国,2001)。
常杰, 陈刚, 葛滢. 1995. 植物结构的分形特征及模拟. 杭州: 杭州大学出版社, 6.
|
李伟成, 潘伯荣, 常杰, 等. 2003. 塔克拉玛干沙漠梭梭地上结构和生物量的自相似模型. 生物数学学报, 18(4): 339-344. |
马克明, 祖元刚. 2000. 植被格局的分形特征. 植物生态学报, 24(1): 111-117. DOI:10.3321/j.issn:1005-264X.2000.01.021 |
潘伯荣, 阎国荣, 真木太一, 等. 1994. 吐鲁番的气候资源及其对种植业的利弊分析. 新疆环境保护, 16(4): 141-151. |
陶玲, 李新荣, 刘新民, 等. 2001. 中国珍稀濒危荒漠植物保护等级的定量研究. 林业科学, 37(1): 52-57. DOI:10.3321/j.issn:1001-7488.2001.01.008 |
邬建国. 2001. 景观生态学——格局、过程、尺度与等级. 北京: 高等教育出版社, 21.
|
Burrough P A. 1981. The factal dimension of landscapes and other environmental data. Nature, 294: 240-242. DOI:10.1038/294240a0 |
Harte J, Kingzig A, Green J. 1999. Self-similarity in the distribution and abundance of Species. Science, 284: 334-336. |
Kaye B K. 1994.徐新阳, 康雁等, 译.分形漫步.沈阳: 东北大学出版社, 67
|
Mandlebrot B. 1979. The fractal geometry of nature. San Francisco: Freeman, 236-237.
|
Nicklas K J. 1986. Computer simulated plant evolution. Science, 54: 78-86. |
Peitgen H O, Jurgens H, Saupe D. 1991. Fractals for the Classroom. New York: Springer-Verlag, 78.
|
Valdez-Cepeda R R, Oliares-Saenz E. 1998. Fractal analysis of Mexico's annual mean yields of maize, bean, wheat and rice. Field Crops Research, 59: 53-62. DOI:10.1016/S0378-4290(98)00108-7 |
Waliszewski P, Konarski J. 2001. Tissue as a self-organizi ng system with fractal dynamics. Cancer and Self-Organization, 28(4): 545-518. |
Waliszewski A, Pattison T R. 1993. Neural Network for Quadratic optimization with bound constrains. IEEE Transactions on Neural Networks, 4(2): 293-304. |