 2006, Vol. 42
2006, Vol. 42文章信息
- 林英华, 孙家宝, 刘海良, 张夫道, 孙龙, 金森.
- Lin Yinghua, Sun Jiabao, Liu Hailiang, Zhang Fudao, Sun Long, Jin Sen.
- 黑龙江帽儿山土壤动物群落组成与多样性分析
- Composition of Soil Fauna Community and its Diversity Analysis at Maoershan Mountains of Heilongjiang Province
- 林业科学, 2006, 42(4): 71-77.
- Scientia Silvae Sinicae, 2006, 42(4): 71-77.
-
文章历史
- 收稿日期:2004-07-30
-
作者相关文章



2. 中国农业科学院土壤肥料研究所 北京 100081;
3. 东北林业大学林学院 哈尔滨 150040;
4. 吉林长白山国家级自然保护区管理局 安图 133613
2. Institute of Soil and Fertilizer, Chinese Academy of Agriculture Sciences Beijing 100081;
3. Forestry College of Northeast Forestry University Harbin 150040;
4. Administration Bureau of National Natural Reserve of Changbaishan Mountains Antu 133613
土壤动物是土壤生态系统的重要组成部分之一,在促进和保护植物群落的次生演替和当地植物物种多样性方面发挥着重要作用,并极大地影响植被的群落结构(de Deyn et al,2003)。森林土壤动物在森林系统中的生态功能性作用已引起学者们的广泛关注,并开展了一系列相关研究,在其生态和功能方面进行了相应的报道(赵小鲁等,1996;殷秀琴等,2001)。
帽儿山地区的植被属长白山植物区系,由地带性顶极植被红松(Pinus koraiensis)阔叶林被人为干扰破坏后,经几个阶段演替而逐步恢复为现在的天然次生林。在演替过程中,土壤动物因其依赖自养和植物控制的微环境,同样经历了与植物演替序列阶段相平行的变化序列,并随着生态系统的变化而变化,形成了与目前帽儿山地区次生林和人工林相对应的土壤动物群落结构。笔者于2002年5—9月,从森林演变的角度,对黑龙江帽儿山地区较为典型的次生林和人工林土壤动物群落进行调查,结果报道如下。
1 研究区概况与方法 1.1 研究区概况研究区位于黑龙江省尚志市帽儿山镇东北林业大学实验林场内的老爷岭森林生态定位站,45°23′—45°27′N,127°34′—127°40′E,海拔353~465 m。该地区属长白山支脉、张广才岭西北部小岭的余脉。该区受欧亚大陆季风气候影响,具有温带季风气候特征。土壤多发育在花岗岩上,地带性土壤为暗棕壤,常见的类型有典型暗棕壤、潜育暗棕壤、白浆化暗棕壤和草甸暗棕壤;非地带性土壤有白浆土、草甸土、沼泽土等。
帽儿山地区的植被,主要以蒙古栎(Quercus mongolica)、白桦(Betula platyphylla)、山杨(Populus davidiana)为主的零星分布以及胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、黄菠萝(Phellodendron amurense)形成的蒙古栎林、山杨林、杨桦椴林、白桦林、胡桃楸林、水曲柳林,草甸和沼泽多种群落类型。
本研究选择4种具有代表性的天然次生红松林(A)、天然次生混交林(B)、人工针叶林(C)、人工混交林(D)为试验样地,分别取样调查。
1.2 研究方法2002年5—9月,在4块样地分3次进行采样。每1种类型样地随机选取3个样方,每个样方内取3个点,每个点重复3次,分别按照枯枝落叶0~5、5~10和10~15 cm分层取样,枯枝落叶取样面积20 cm×20 cm。
受条件限制,本次调查仅采用手拣和改良Tullgren法分离提取土壤动物并分别鉴定统计数量,以干生土壤动物为主(尹文英等,1998)。体型大小依据在食物分解过程中的作用(Swift et al., 1979)进行划分。
1.3 数据处理群落多样性指数采用香农-威纳多样性指数(Shannon-Weaner index)、Pielou指数和辛普森优势度指数(Simpson index),即
群落相似性采用Jaccard(q)指数、Gower系数(W(j, r)),即q=c/(a+b-c),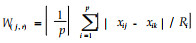
采用非线性统计分析方法Kruskal-Wallis检验法分析不同林型之间土壤动物的差别。各类群数量等级划分:个体数量占全部捕获量10%以上为优势类群,介于1%~10%之间的为常见类群,介于0.1%~1%为稀有类群,0.1%以下的为极稀有类群。
2 结果与分析 2.1 土壤动物群落特征在天然次生针叶林(A)、天然次生混交林(B)、人工针叶林(C)、人工混交林(D)4块样地内,3次(2002-05—2002-09)共获得土壤动物30 045只(未鉴定个体24只),隶属3门10纲21目3亚目56科,其中土层(0~15 cm)7 631只,隶属3门10纲21目3亚目41科,森林凋落物层22 390只,隶属3门10纲21目3亚目50科,见表 1。
|
|
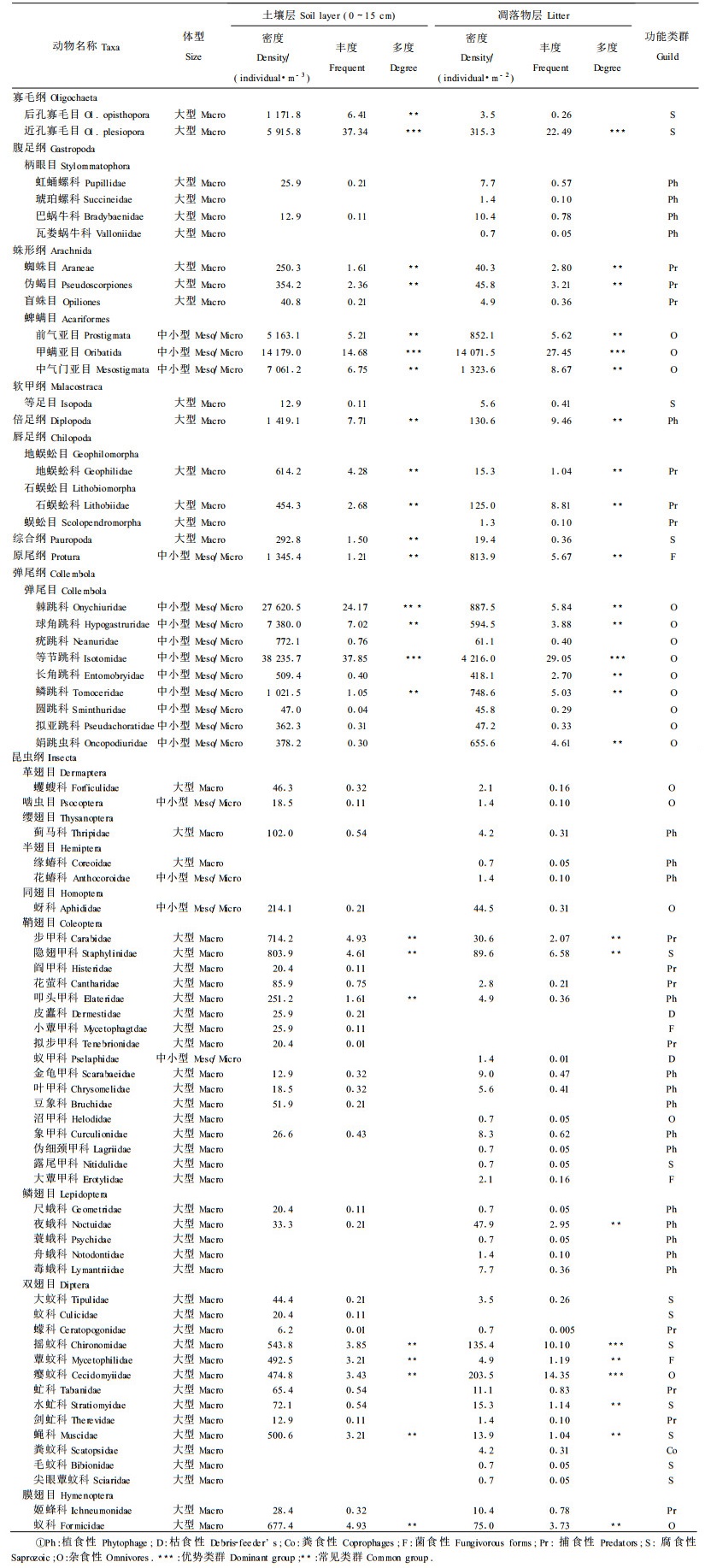
从表 1中可以看出,分布土壤0~15 cm的大型土壤动物40类,优势类群为近孔寡毛目;常见类群15类:综合纲、蜘蛛目、叩头甲、伪蝎目、石蜈蚣目、蕈蚊科、蝇科、瘿蚊科、摇蚊科、地蜈蚣科、隐翅甲科、步甲科、蚁科、后孔寡毛目、倍足纲。中、小型土壤动物15类,优势类群3类:蜱螨目甲螨亚目、棘跳科、等节跳科;常见类群7类:疣跳科、鳞跳科、原尾纲、线虫动物门、蜱螨目前气门亚目、蜱螨目中气门亚目、球角跳科。分布在森林凋落物层的大型土壤动物优势类群为摇蚊科、瘿蚊科、近孔寡毛目;常见类群为地蜈蚣科、蝇科、水虻科、蕈蚊科、步甲科、蜘蛛目、夜蛾科、伪蝎目、蚁科、隐翅甲科、石蜈蚣目、唇足纲。中、小型土壤动物优势类群为蜱螨目甲螨亚目、等节跳科,常见类群为长角跳科、球角跳科、娟跳虫科、鳞跳科、蜱螨目前气门亚目、原尾纲、棘跳科、蜱螨目中气门亚目。这些土壤动物构成了帽儿山森林土壤动物的主要类群,它们对该森林土壤环境具有较强的适应能力。
分布在土壤层和森林凋落物层的稀有类群(占总个体数0.1%~1%):大型土壤动物分别有2 3类和25类;中小型土壤动物分别为5类和6类。极稀有类群(占总个体数0.1%以下)分别有2类和10类,中小型土壤动物中仅土壤层1类,这与它们对所处的森林环境的变化较为敏感有关,在某一时期土壤条件适宜时,其种群数量才会逐渐增加,并成为某一时期的常见类群。
2.2 土壤动物群落动态变化 2.2.1 土壤动物的林型分布不同林型土壤动物群落,即水平分布(表 2),4种林型共有类群35类,仅分布在1种林型内的类群有18类,其中A林型7类、B林型6类、C林型0类、D林型5类,不同林型之间土壤动物个体数量和类群差异明显(H′number=7.81,α>0.10; H′group=10.19,α>0.05);土壤层个体数量和类群数排序依次均为D(63.90×104·m-3, 56类)>A(44.28×104·m-3, 52类)>B(22.24 ×104·m-3, 49类)>C(13.13×104·m-3, 41类)。
同一林型中土壤动物个体和类群数垂直分布现象显著。土壤动物个体数量一般表现为0~5 c m最大,10~15 cm最少;类群均随着土壤剖面层次的加深而递减。在调查中同时发现,土壤动物大量集聚在森林凋落物层,尤其是人工林纯林凋落物层分布的土壤动物最多,高达总数的84.56%,这是因为凋落物分解释放出丰富有机物,导致土壤动物大量集聚;而混交林中,由于阔叶林凋落物分解的速度高于针叶林的分解速度,因而土壤表层(0~5 cm)所分布的土壤动物明显地多于针叶纯林,因此土壤动物类群和个体数较为丰富。
2.2.2 土壤动物群落月变化整体而言,不同月份土壤动物总数量和类群总数变化趋势不一致(表 2),其中森林凋落物层的土壤动物个体数量依次是9月份(9.74×104·m-2)>5月份(6.07×104 ·m -2)>7月份(5.46×104·m-2),类群数依次9月份(50类)>7月份(47类)>5月份(41类);土壤层的土壤动物数量月变化与森林凋落物层一致,其个体数量依次是9月份(69.83×104·m-3)>5月份(56.55×104·m-3)>7月份(38.44×104·m-3),类群数依次9月份(57类)>7月份(55类)>5月份(47类)。
|
|
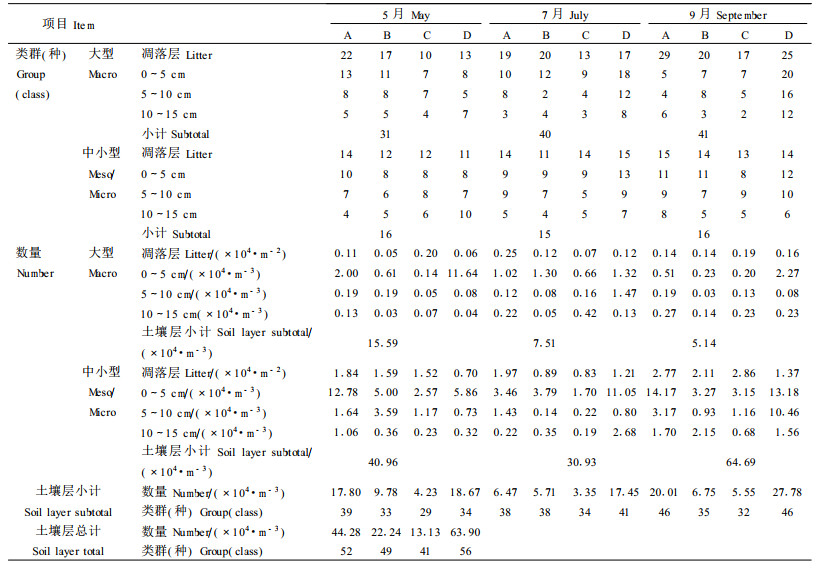
不同体型的土壤动物数量和类群数分布与土壤动物总数量和类群总数分布略有差异,其中凋落物层大型土壤动物数量和类群数随月份呈递增趋势,即9月份(0.63×104·m-2,35类)>7月份(0.56×104·m-2,32类)>5月份(0.42×104·m-2,27类);中小型土壤动物类群数则为9月份(15类)=7月份(15类)>5月份(14类);中小型土壤动物数量为9月份(9.11×104·m-2)>5月份(5.65×104·m-2)>7月份(4.90×104·m-2)。土壤层的大型动物则5月份(15.59×104·m-3)>7月份(7.51×104·m-3)>9月份(5.14×104·m-3),类群数则为9月份(30类)=7月份(30类)>5月份(24类);中小型土壤动物数量为9月份(64.69×1 04·m-3)>5月份(40.96×104·m-3)>7月份(30.93×104·m -3),中小型土壤动物类群数3个月份均为14类。
2.2.3 土壤动物群落多样性与均匀性土壤动物多样性与均匀性如表 3所示,其中多样性指数(H′)最高值和最低值均出现在9月份;林型A、B、C 5月份土壤动物的多样性均最高;林型D则9月份土壤动物的多样性最高。
|
|

Pielou指数(Js)和辛普森优势度指数(C)总体水平的月变化与多样性指数相同,但各林型月份变化与多样性变化有所差异,如林型B中,5月份土壤动物的均匀性最低,7月份则最高;林型C中,5月份土壤动物的多样性最高,而9月份最低;林型D中,9月份土壤动物的多样性最高,而5月份最低。土壤动物均匀度的最大值通常出现在种类和数量较少的林型;均匀度的最小值出现在种类和数量较多的林型。说明在林型单一或条件严酷的环境中通常土壤动物稀有种类较少,优势类群和常见类群占优势,因此种间差异较小,均匀度较高,优势现象不明显。从群落多样性看,各林型内香农-威纳指数和均匀度指数变化较大,群落的各项指标季节变化大,稳定性较差。
由于H′指数为Js和C指数的函数,反映系统信息的大小,具有可累加性,将每种林型不同月份的H′指数进行累加,其大小依次为D(9.617 6)>C(9.073 5)>A(8.5 22 8)>B(7.804 7),表明次生林土壤动物群落多样性低于人工林,其林内小环境和土壤环境条件相对较差。
2.2.4 土壤动物群落相似性在4种林型中,A与B之间的Jaccard(q)指数明显低于其他林型之间的系数,而B与C之间的Gower系数明显低于其他林型之间的系数。如表 3,其中Jaccard(q)系数越小,表明其相似程度越低;Gower系数则相反,其值越高,相似程度越低,表明不同林型土壤动物群落的组成具有较高的异质性,反映出不同林型对土壤生态环境、进而对土壤动物群落产生影响。从2种分析指标来看,由于4种林型共有类群较多,少数共有类群相应个体相差较大,Jaccard(q)指数、Gower系数较高。
3 结论与讨论土壤动物群落特征与土壤的各种特性,如水、pH值、结构、质地、土壤有机质含量以及环境中植被盖度和凋落物年归还量等与土壤动物均有密切关系,并随着生态系统的异常变化和植物诱导的自发变化而变化。
总体而言,天然次生林土壤动物个体总数(66.52×104·m-3)低于人工林的个体总数(78.03×104·m-3),但类群数(63)却高于人工林(57);在天然次生林中,次生针叶林土壤动物数量和类群数明显高于次生混交林,而人工林则相反,在某种意义上反映出天然次生林与人工林生态环境对土壤动物的不同影响,反映了帽儿山森林土壤动物的组成改变与森林生态环境变迁之间的联系,其中人工林土壤动物个体数量和类群数的变化与张雪萍(1995)的研究结果相符。
5、7、9月份采集的土壤动物数量和类群分析结果表明,3个月份土壤动物的个体数量和类群数分布都存在着不均衡的现象,总的趋势分别是9月>5月>7月、9月(57类)>7月(55类)>5月(47类),表明帽儿山土壤动物数量和类群的波动除了与生态因子中的温度、日照、湿度、降水量相关以外(于长福等,1985),还可能与盖度、凋落物厚度等有关,Mikola等(2001)已证实了落叶和部分落叶植物能够改变土壤生物的特性,并且植物叶片的数量与特性能影响土壤生物的种群和类群动态(Aubert et al., 2004),而不同植物的凋落物可能对大型土壤动物产生影响(Wardle et al., 2002),这与本研究凋落物的厚度对土壤动物个体数量和类群影响较大相一致。
在众多表征群落多样性的指数中,香农-威纳多样性指数和辛普森多样性指数是应用最广泛的2个指数,一些学者们将其应用于群落多样性的分析,但由于土壤动物受分析的限制,许多土壤动物群落等级属性表现非常明显,因此在分析比较群落的多样性时,生物的等级应予以考虑,Pielou(马克平等,1994)提出了信息多样性指数测度等级多样性方法,即对于科、属、种3个等级的多样性,有H′(SGF)=H′(F)+H′F(G)+H′FG(S),其中H′(F)为分类单元的多样性;H′F(G)为以F内G为分类单元对各F的多样性的加权平均;H′FG(S)为以G内S如分类单元的多样性的加权平均,它表明H′(SGF)在相同的群落内,H′(F)或H′F(G)值更大的类群具有更高的复杂性。依据该指数计算4种林型土壤动物多样性值依次为3.005 2、2.878 4、3.151 7和3.499 3,其值整体低于香农-威纳多样性指数(分别为3.920 0、3.630 5、3.990 7和4.27 0 1),但是其变化趋势与香农-威纳多样性指数变化趋势一致,因此采用2种分析方法反映出相同的结论。
Jaccard(q)指数和Gower系数是群落相似性分析中较常用的分析方法。Jaccard(q)指数适于某个类群存在或不存在的二元数据,而后者是作为衡量2个实体(属性)间相异性的指标,前者数值大小反映了群落之间的相似程度,数值越大,表明2个群落之间越相似;后者则反映群落之间差异程度,数值越小,表明2个群落之间的相似程度越大。采用相似性和相异性2种指标对4种林型土壤动物群落的相似性进行分析,能更好地反映出群落之间的相互关系。但分析中发现,2种指数表征的相似程度不一致,Jaccard(q)指数反映出B与D之间最相似,其次是C与D之间,A与B之间相似性最差;Gower系数则是B与C最相似,其次是A与C,而B与D之间相似性最差,两者误差的产生及原因,有待于今后进一步研究。
由于受条件限制,本研究仅采用手拣法和大型改良干漏斗对帽儿山的土壤动物进行采集,并以干性土壤动物为主。由于土壤动物对环境因子较为敏感,一些敏感性种类仅在某一时期土壤条件适宜时,种群数量才逐渐增加,并成为某一时期的常见的类群,这与以往学者对帽儿山土壤动物群落研究结果基本一致。
马克平, 刘玉明. 1994. 生物群落多样性的测量方法Ⅰ: α多样性的测量方法(下). 生物多样性, 2(4): 231-239. DOI:10.3321/j.issn:1005-0094.1994.04.009 |
殷秀琴. 2001. 东北森林土壤动物研究. 长春: 东北师范大学出版社.
|
于长福, 赵克尊. 1985. 老爷岭生态实验站的土壤动物初步调查研究. 东北林业大学学报, 13(3): 128-134. |
张雪萍. 1995. 帽儿山针叶林和针阔混交林土壤动物对比研究Ⅰ:区系组成与特征. 哈东滨师范大学自然科学学报, 11(2): 94-99. |
赵小鲁, 谢丙庚. 1996. 动物生态地理研究——陈鹏教授等论文集. 成都: 成都地图出版社.
|
尹文英. 1998. 中国土壤动物检索图鉴. 北京: 科学出版社.
|
Aubert M, Hedde M, Decaëns T, et al. 2004. Effects of tree canopy composition on earthworms and other macro-invertebrates in beech forests of Upper Norman dy (France). Pedobiologia, 47(5-6): 904-912. |
de Deyn G B, Raaijmakers C E, Zoomer H R, et al. 2003. Soil invertebrate fauna enhances grassland succession and diversity. Nature, 422(17): 711-713. |
Mikola J, Yeates G W, Wardle D A, et al. 2001. Response of soil food-web structure to defoliation of different plant species combinations in an experimental grassland community. Soil Biology and Biochemical, 33: 205-214. DOI:10.1016/S0038-0717(00)00131-0 |
Swift M J, Heal O W, Anderson J M. 1979. Decomposition in terrestrial ecosystems. Univ Calif Press Berkeley
|
Wardle D A, Bonner K I, Barker G M. 2002. Linkages between plant litter decomposition, litter quality, and vegetation responses to herbivores. Functional Ecology, 16: 585-595. DOI:10.1046/j.1365-2435.2002.00659.x |