 2006, Vol. 42
2006, Vol. 42文章信息
- 杨锦昌, 许煌灿, 尹光天, 冯昌林, 卢立华.
- Yang Jinchang, Xu Huangcan, Yin Guangtian, Feng Changlin, Lu Lihua.
- 黄藤人工林密度效应
- The Density Effect of Daemonorops margaritae Plantation
- 林业科学, 2006, 42(4): 57-61.
- Scientia Silvae Sinicae, 2006, 42(4): 57-61.
-
文章历史
- 收稿日期:2003-09-28
-
作者相关文章


2. 中国林业科学研究院热带林业实验中心 凭祥 532600
2. Experimental Center of Tropical Forestry, CAF Pingxiang 532600
造林密度是影响林分生长与收获的重要因子, 也是提高人工林生产力的关键措施。因而密度管理不仅是林业生产需要解决的关键技术之一, 也成为林业科学研究的热点。许多学者对杉木(Cunninghamia lanceolata)、马尾松(Pinus massoniana)、毛竹(Phyllostachys pubescens)、杨树(Populus spp.)、长白山落叶松(Larix olgensis)等人工林进行深入研究, 取得了可喜的成果, 并有效地指导了林业生产(叶功富等, 1995;童书振等, 2002;刘金福等, 1998;洪伟等, 1998;沈作奎等, 2002;吴春芳, 1998;张怡春等, 2000)。国内外也对棕榈藤(Rattan)人工林造林密度开展了大量试验(Wong et al., 1985;Chin, 1989;Cecilia et al., 1992;Nur Supardi et al., 1992;许煌灿等, 1994;Rao et al., 1997), 但尚缺乏定量描述和系统分析, 对不同藤种采用哪种造林密度获得理想效果则鲜有报道。为此, 以黄藤(Daemonorops margariae)人工林为对象, 研究一定立地条件下不同造林密度对棕榈藤林分动态及生长规律的影响, 探讨黄藤人工林的密度效应, 为藤林经营管理提供科学依据。
1 试验地概况黄藤试验地位于广西凭祥市热带林业实验中心英阳试验站, 22°07′ N, 106°44′ E, 属南亚热带季风气候, 干湿季节交替明显。年平均气温21.6 ℃, 最冷的1月份平均气温13.3 ℃, 极端最低气温0.1 ℃, 最热的7月份平均气温27.7 ℃, 极端最高气温37.9 ℃, ≥10 ℃积温7 596 ℃, 年均降水量1 388 mm, 年蒸发量1 275 mm。试验地上层树种为马尾松(P.massoniana), 1984年造林, 密度1 650株·hm-2; 试验地海拔320~360 m; 土壤为花岗岩发育而成的砖红壤性红壤, 土层厚度150 cm以上, 表土层腐殖质含量26.1~40.2 g·kg-1, 含氮量1.024~1.374 g·kg-1, 速效磷含量13.59~18.01 mg·kg-1, 速效钾含量17.5~30.5 mg·kg-1。
2 材料与方法 2.1 造林措施种植前清理林下杂草和灌木, 适当疏伐林木, 控制上层林分郁闭度约0.5;穴状整地, 种植穴规格为50 cm×50 cm×30 cm。用生长整齐的藤苗, 单株种植, 造林后3个月检查成活率并加以补植, 每年抚育2次持续到第3年。
2.2 试验安排采用随机区组设计, 安排4个区组, 4种密度处理, 即1 m×2 m(D), 1 m×3 m(C), 2 m×3 m(B)和3 m×3 m(A), 共16个小区。试验小区为长方形, 每小区面积600 m2。每个小区的四周设置2行相同密度的保护带。
2.3 数据采集和处理造林后第2年只测定苗高, 第3~5年测定各小区藤丛的萌蘖数和植株长; 当年龄达14年时, 调查各小区死亡藤丛数以此计算自疏率, 并采用随机取样方法抽取30个藤丛测定其株数、长度; 同时采收长度4 m以上的藤株, 测量其2 m处的直径、鲜藤质量。数据的处理和分析在计算机上利用SPSS统计软件进行。
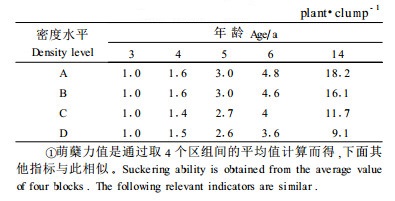
3 结果与分析 3.1 密度对萌蘖力的影响萌蘖既是棕榈藤重要的生物学特性, 又是棕榈藤的重要生长指标, 它导致植株数量不断增加, 进而增加藤丛的总茎长(曾炳山等, 1993;许煌灿等, 1994)。萌蘖力可用藤丛的平均株数来表示。表 1反映了萌蘖力在不同年龄时随密度的变化规律:藤丛的平均株数因年龄的推移而不断增加, 随密度的增大而减少。在3年时不同密度间的藤株均不产生萌蘖; 到了4年, 藤株才开始萌蘖, 但不同密度间差异不大; 随后藤株萌蘖力不断增强, 不同密度处理间的差异逐步拉大。方差分析结果(表 2)表明, 4~5年时区组间有显著差异而密度间无显著差异; 6年时, 不同密度对萌蘖力的影响达到显著水平, 经Q检验, A与D之间差异显著; 14年时, 密度间的差异达到极显著, 其中A、B与C、D之间差异均达到极显著水平, 其他密度间无显著差异。不同密度间的萌蘖力的大小依次为A>B>C>D。
|
|
|
|
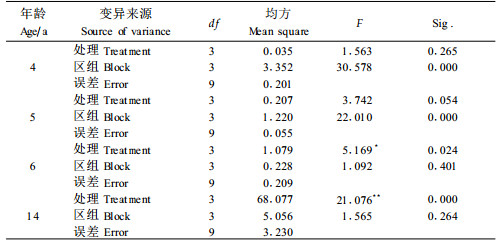
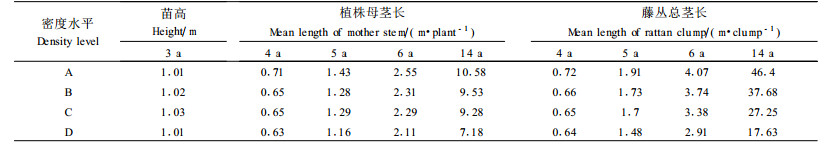
随着年龄的增加, 藤丛不仅横向萌蘖扩展, 而且纵向攀缘伸长, 导致藤丛茎长不断发生变化。藤林年龄为3年时, 绝大部分植株还没有形成藤茎, 故只测定苗高, 而后开始产生藤茎。从表 3可看出, 藤林年龄为3年时, 不同处理间的苗高几乎没有差异, 此时, 密度对苗木生长几乎没有影响; 随着年龄的增大, 植株母茎长及藤丛总茎长不断增加, 在不同密度间的差异也逐步增大。方差分析结果(表 5)表明, 4~6年时植株母茎长和藤丛总茎长在区组间存在极显著差异, 而在密度处理间无显著差异; 14年时, 密度对植株母茎长仍无显著影响, 但对藤丛总茎长的影响达到极显著水平。通过Q检验得出:14年时藤丛总茎长在密度A与C、D之间分别存在显著和极显著差异。
|
|
|
|

|
|
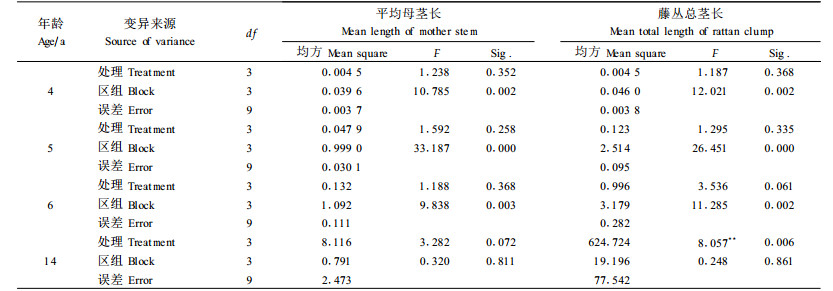
由表 4可知, 对于不同密度的试验林, 其自疏率有较大的变化。方差分析结果(表 8)表明, 不同密度对自疏率产生极显著影响。经Q检验, A与C之间有显著差异, 而A与D之间则有极显著差异。由此可知, 密度对藤林自然稀疏有着重要的影响, 密度越大, 自疏率也越大。
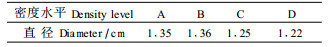
3.4 密度对直径的影响总体上, 直径随着密度的增加而递减。当造林密度减少到1 111株·hm-2时, 直径几乎没有变化, 甚至略微变小(表 6)。A与B, C与D之间直径差异较小; 而A、B与C、D之间差异较大。通过方差分析(表 8)可知, 密度对林分直径产生显著影响; 多重比较结果表明, 林分直径在密度B与D之间存在显著差异, 而其他密度水平间无显著差异。
|
|
|
|

|
|
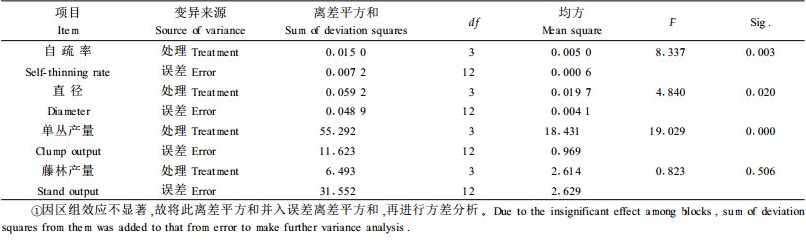
自疏率、萌蘖力、平均长度和直径最终共同影响到藤林的产量, 本文用鲜藤质量来表示其产量。根据14年时的采收结果(表 7)可知, 单丛产量随密度的增加而不断减少, 而藤林产量随着密度的增加先增加后逐步减少, 尤其在B与C之间产量差异特别明显。经方差分析(表 8), 密度不同造成了单丛产量的显著差异; 通过Q检验表明:A与C、D之间差异均达到极显著水平, 而B与C、D之间分别产生显著和极显著差异; 而不同密度对藤林总产量则无显著差异。
4 结论与讨论1) 黄藤的萌蘖力随年龄的增加而增大, 随造林密度的增加而递减。当藤林年龄≤5年时, 不同密度的萌蘖力在不同密度间无明显差别; 而后, 随年龄不断增加, 不同密度处理间的差异迅速增大, 以致达到显著或极显著水平; 各密度间的萌蘖力大小依次为D>C>B>A。藤株仅在年龄≥4年, 即造林3年之后才开始萌蘖, 说明黄藤植株需要达一定年龄才抽茎萌蘖, 但其萌蘖的初始时间与曾炳山等(1993)的研究结果不太一致, 这可能与造林时所选藤苗和立地条件有关。在开始萌蘖的初期, 萌蘖力较弱, 不同密度的萌蘖数均较小; 到了后期萌蘖力逐步增强, 藤丛的平均株数也不断增加, 这与许煌灿等(1994)的研究结论相似。萌蘖力在后期的显著差异表明:密度越大, 藤林越早郁闭, 植株间的竞争也越激烈, 从而导致萌蘖数的减少。
2) 母茎长和藤丛平均总长均在一定程度上受造林密度的影响, 随密度的增加而递减。然而, 在年龄≤6年时, 密度不论对苗高, 还是对母茎长或藤丛平均总长均影响不大, 不同密度间无显著差异; 14年时调查结果表明, 密度对植株母茎长无显著影响, 但藤丛总茎长产生极显著差异。在相同密度条件下, 总茎长均大于母茎长, 二者间的差异随年龄的增加而增大, 说明密度对藤丛平均总长的影响大于对母茎长的影响。这是由于藤丛平均总长是由藤丛内各藤株茎长组成(曾炳山等, 1993;许煌灿等, 1994), 它与萌蘖数联系紧密, 而萌蘖数又受密度的影响, 从而产生累积效应; 相反, 母茎长则与萌蘖数关系不大。
3) 造林密度对藤林自然稀疏有着重要的影响, 自疏率随密度的增加而递增, 不同密度间自疏率的大小分别是D>C>B>A。这种变化规律是林分自然生长过程的一大特征(唐守正, 1993), 反映了不同密度条件下营养空间的动态变化及藤丛间的竞争程度。虽然黄藤与一般林木的生态生物学特性差异较大, 但密度对黄藤自稀率的影响与其他林木(童书振等, 2002;江希钿等, 2001)基本相似。
4) 直径大致随着密度的增加而递减; 但造林密度减小或增大至一定程度时, 密度对直径所产生效应并不明显, 过密或过疏均不利于获得理想的效果。密度为1 111株·hm-2时的直径略小于密度为1 667株·hm-2时直径, 这可能与密度较稀时由于藤丛萌蘖数较多、植株间的竞争较激烈有关; 当然, 这个解释有待于进一步验证。由于棕榈藤的藤茎粗度变化不均, 通常基部较粗, 向上生长渐细, 约至2~3 m处变化很小(蔡则谟等, 2003), 因而确定藤株直径时在距基部约2 m处测量而不是在1.3 m处。
5) 单丛平均产量随造林密度的增加而递减, 并以密度B与C间的单丛产量差异最大。除密度A与B及C与D之间无显著差异外, 其他密度间均达到显著或极显著差异。与前面几个测量指标相比, 密度对其单丛平均产量的影响最大。而藤林总产量则随着密度的增大先增加后逐步减少, 不同密度间无显著差异, 但以密度每公顷3 333株的总产量最高。藤林的单丛产量和总产量随密度的变化规律与前人提出的林分密度效应相符(刘金福等, 1998), 两者是上述几个测量因子的共同作用的表现, 也是造林密度效应的综合反映。
虽然密度越小, 单丛产量越高, 但总产量却不一定越大, 林分密度与藤林总产量的关系呈二次效应模型(洪伟等, 1996;陈辉等, 1992), 因而选择合适的造林密度对于获得最高的藤产量至关重要。根据研究结果, 藤丛的总产量分别为C>D>B>A, 因此, 单从产量考虑, 造林时应采用密度C和D。然而D密度太大, 无法对林分进行有效的管理和采收(Nur Supardi et al., 1992), 而且造林成本偏大; 密度B的总产量尽管比C和D都低, 但株行距均比较大, 便于管理, 因而在实际上可采用密度B; 而密度A太小, 不能充分利用林地。综上所述, 进行营建黄藤人工林时, 可根据上层林分的实际情况采用3 m×1 m或3 m×2 m密度, 从而既获得较高的藤产量, 又方便藤林的管理。
需要指出的是本试验仅探讨了林分密度单个因素对黄藤生长的影响, 实际上, 立地条件、上层林分郁闭等也是影响藤林生长的重要因素(许煌灿等, 1994;Manokaran, 1984), 并往往与密度起交互作用共同决定藤林的生长与收获, 因此, 多因子对藤林生长的影响有待于今后加强研究。
蔡则谟, 许煌灿, 尹光天, 等. 2003. 棕榈藤利用的研究与进展. 林业科学研究, 16(4): 479-487. DOI:10.3321/j.issn:1001-1498.2003.04.018 |
陈辉, 何宗明, 洪伟. 1992. 杉木人工林密度效应模型研究. 福建林学院学报, 12(3): 277-282. |
洪伟, 郑郁善, 邱尔发. 1998. 毛竹丰产林密度效应研究. 林业科学, 34(专): 1-4. |
洪伟, 吴承祯. 1996. 闽北杉木人工林密度控制连续状态的动态规划研究. 福建林学院学报, 16(1): 1-4. |
江希钿, 王素萍, 杨锦昌. 2001. 马尾松人工林种群自然稀疏模型的研究. 热带亚热带植物学报, 9(4): 290-295. |
刘金福, 洪伟, 林芳. 1998. 马尾松人工林密度效应模型研究. 西南林学院学报, 18(3): 148-152. |
沈作奎, 徐伟声, 蒋剑波. 2002. 日本落叶松人工林密度效应的研究. 湖北民族学院学报:自然科学版, 20(4): 11-13. |
童书振, 盛炜彤, 张建国. 2002. 杉木林分密度效应研究. 林业科学研究, 15(1): 66-75. DOI:10.3321/j.issn:1001-1498.2002.01.011 |
唐守正. 1993. 同龄纯林自疏规律的研究. 林业科学, 29(3): 235-241. DOI:10.3321/j.issn:1001-1498.1993.03.005 |
吴春芳, 王玉亭, 冯俊一, 等. 1998. 北京杨速生丰产林密度效应. 青海农林科技, 2: 18-20. |
许煌灿, 尹光天, 曾炳山. 1994. 棕榈藤的研究. 广州: 广东科技出版社, 60-66.
|
叶功富, 林武星, 张水松, 等. 1995. 不同密度管理措施对杉木林分的生长、生态效应的研究. 福建林业科技, 22(3): 1-8. |
曾炳山, 许煌灿, 尹光天, 等. 1993. 黄藤藤丛结构和生长的研究. 林业科学研究, 6(4): 414-423. DOI:10.3321/j.issn:1001-1498.1993.04.001 |
张怡春, 兰湛, 王丽艳, 等. 2000. 长白落叶松人工林密度效应模型的研究. 吉林林业科技, 29(2): 21-22, 61. |
Cecilia M L, Faustina C B, Rowena V P.1992. A Beginner's Source book on Philippine Rattan.Philippine Council for Agriculture, Forestry and Natural Resources Research and Development, 12-28
|
Chin T H. 1989.Rattan planting. Leaflet No. 125, the Department of Agriculture, Sarawak, 18-41
|
Manokaran N. 1984.Biological and ecological considerations pertinent to the silviculture of rattan//Proceedings of the rattan seminar, Kuala Lumpur, Malaysia, 2-4 Oct. The Rattan Information Center, FRIM, 95-105
|
Rao A N, Rao V R.1997.Rattan Taxonomy, Ecology Silviculture, Conservation, Genetic improvement and Biotechnology.IPGRI-APO, Serdang, Malaysia, 64-69
|
Nur Supardi M N, Aminuddin M. 1992.Planting systems for large-diameter canes//Razali W, Dransfield J, Manokaran N. A guide to the cultivation of rattans.Malaysian Forest Records No. 35. Forest Research Institute Malaysia, 89-98
|
Wong K M, Manokaran N.1985. Proceedings of the Rattan Seminar. October 2-4, 1984, Kuala Lumpur, Malaysia, RIC
|