 2006, Vol. 42
2006, Vol. 42文章信息
- 郑国生, 何秀丽.
- Zheng Guosheng, He Xiuli.
- 夏季遮荫改善大田牡丹叶片光合功能的研究
- Studies on the Photosynthetic Improvement in the Leaves of Field Tree Peony through Shading Treatment in Summer
- 林业科学, 2006, 42(4): 27-32.
- Scientia Silvae Sinicae, 2006, 42(4): 27-32.
-
文章历史
- 收稿日期:2004-10-08
-
作者相关文章


2. 中国农业大学农学与生物技术学院 北京 100094;
3. 山东农业大学生命科学学院 泰安 271018
2. College of Agronomy and Biotechnology, China Agricultural University Beijing 100094;
3. College of Life Science, Shandong Agricultural University Tai'an 271018
光合机构吸收的光能超过光合作用的利用量时光合活性降低, 即出现光抑制现象。过剩的光能通过多种途径在植物体内产生有毒的活性氧, 导致活性氧伤害(徐志防等, 1998);光饱和解联的电子传递活性受抑制(付振书等, 2005), 光合碳代谢酶活性下降(江力等, 2000);在有氧条件下光抑制会引起多种膜蛋白及膜脂破坏(郭晓荣等, 2004)。关于植物的光抑制, 近年来研究较多, 对光抑制的机理研究也取得了很大进展。研究(Barber, 1995;Critchley et al., 1994)表明, 强光下植物发生光抑制的主要部位是光系统Ⅱ(PSⅡ); 光合作用日变化能够反映光合作用一天的变化情况, 是研究光抑制的有效指标。对小麦(Triticum aestivum)(郭连旺等, 1996)、水稻(Oryza sativa)(李霞等, 2002)、生姜(Zingiber officinale)(张振贤等, 2000)等作物光合作用日变化的研究证明, 光抑制是普遍存在的现象。适度遮荫可以使净光合速率(Pn)增加, 气孔导度(Gs)增大, 光能和水分利用效率提高(刘贤赵等, 2002;艾希珍等, 2000)。
牡丹(Paeonia suffruticosa)原产我国西北部, 喜凉恶热, 宜燥惧湿, 夏季高温、强光下, 叶片常常卷曲枯焦, 影响生长(蓝宝卿等, 2002)。陈向明等(2001)对保护地栽培牡丹的光合特性进行了研究, 而关于大田牡丹叶片的光合特性、光抑制以及遮荫对其影响未见报道。本文对自然光下大田牡丹在生长最适宜的春季、高温强光下生长受抑制的夏季, 以及夏季遮荫处理下牡丹叶片的光合作用日变化进行了研究, 旨在探讨大田牡丹在夏季高温强光下光合作用的变化规律, 是否存在或发生了光抑制, 以及夏季遮荫减轻或克服光抑制的机理, 为改进大田牡丹的栽培技术提供理论依据。
1 材料与方法 1.1 供试材料及处理试验于2004年在山东泰安牡丹苗圃试验田进行, 供试品种为‘大胡红’(P. suffruticosa cv.‘Dahuhong’)。精选生长一致、生长势强的牡丹60株, 于6月30日用一层纱网进行棚式遮荫(遮光50%)处理, 以自然光下生长的60株为对照。春季测定为4月28日、5月8日和5月18日, 日平均气温分别为23、25和26 ℃; 夏季测定为7月16日、7月26日和8月6日, 日平均气温分别为32、34和35 ℃。其他管理按常规进行。
1.2 牡丹叶片CO2气体交换参数的测定用英国PP Systems科学仪器公司生产的TPS-1型便携式光合测定系统, 在晴天选取充分受光、叶位一致的叶片测定光合速率(Pn), 仪器同时读出气孔导度(Gs)、细胞间隙CO2浓度(Ci)、蒸腾速率(Tr)、光合有效辐射(PAR)、叶温(tl)、气温(ta)、大气CO2浓度(Ca), 试验结果为6次测定的平均值。光合日变化测定从8:00—16:00, 每隔1 h测定1次Pn, 每次测定固定的10个叶片, 在30 min内完成, 取其平均值。通过纱布遮光逐步改变PAR, 测定牡丹叶片的Pn, 制作PAR-Pn响应曲线, 并对低于200 μmol·m-2 s-1光强下的Pn和PAR进行线性回归计算量子产额, 其斜率为表观量子效率(AQY)(许大全等, 1987)。在饱和光强下, 测定牡丹叶片的Pn, 对低于250 μL·L-1下的Ci和对应的Pn进行线性回归, 求得初始斜率为RuBP羧化效率(CE)(张振贤等, 2000)。按Berry和Downton(1982)的方法计算气孔限制值(Ls)。
1.3 叶绿素荧光参数的测定测定光合作用日变化的同时, 用英国Hansatech科学仪器公司生产的植物效率仪(HPEA)进行牡丹叶片叶绿素荧光参数日变化的测定, 包括初始荧光(Fo)、最大荧光(Fm)和PSⅡ最大光化学效率(Fv/Fm), 每隔1 h测定1次, 每次测定6个叶片, 取其平均值, 测定前将牡丹叶片暗适应15 min。
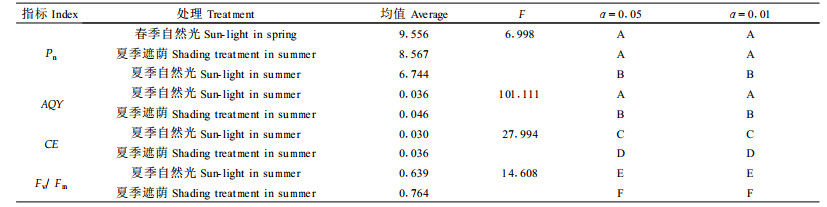
2 结果与分析 2.1 大田牡丹叶片净光合速率(Pn)的日变化从图 1 b(10:00左右测定)可以看出, 大田牡丹叶片的饱和光强约为750 μmol·m-2 s-1, 计算得光补偿点约为18 μmol·m-2 s-1。从图 1 a、c、d看出, 自然光下春季牡丹叶片的Pn、PAR和ta日变化均呈“单峰”曲线, 13:00左右PAR(1 000 μmol·m-2 s-1, 高出饱和光强较少)和ta达到最大值(28.5 ℃, 处于植物叶片光合作用的最适温度范围)时, Pn只有少许降低。自然光下夏季Pn日变化呈“双峰”曲线, 峰值分别出现在10:00和15:00左右, 第2峰值低于第1峰值, 13:00左右Pn降低很多, 此时, PAR最高值为1 430 μmol·m-2 s-1, 高出饱和光强很多, 而且持续时间长, 受强光的影响, 最高气温达37.5 ℃, 高出光合作用的适宜温度很多, 因而出现了严重的光合“午休”; 与春季Pn日变化比较, 中午13:00约降低了57.26%。夏季遮荫处理后, Pn日变化呈“单峰”曲线, 与春季自然光下Pn日变化趋势相似(图 1 a); 中午PAR仅在735 μmol·m-2 s-1左右(图 1 c), 接近牡丹叶片的饱和光强, ta也略有降低(图 1 d)达36 ℃, 但由于受整个大气环境温度的影响降低幅度不大; Pn值明显比夏季自然光下高(中午13:00时约高52.38%), 但比春季低(中午13:00时约低10.25%)。LSD检验(表 2)表明, 春季自然光与夏季自然光下Pn日变化在P < 0.01水平上差异极显著, 夏季遮荫处理与夏季自然光下Pn日变化在P < 0.05水平上差异显著, 而春季自然光与夏季遮荫处理差异不显著。这也说明, 夏季自然光下限制牡丹叶片光合作用的重要因子是强光, 适度遮荫可以明显地改善和克服光合午休, 促进光合作用。

|
图 1 大田牡丹叶片净光合速率对净光合速率(Pn)(a)、光强的响应曲线(b)和光合有效辐射(PAR)(c)、气温(ta)(d)的日变化 Fig. 1 Photosynthesis-light response curves and diurnal variation of net photosynthetic rate (Pn) and photosynthetic active radiation (PAR) and air temperature (ta) in the leaves of field Tree Peony |
结合表 1可以看出, 12:00与8:00相比, 夏季自然光下生长的牡丹叶片, 随着光强和气温的升高, Gs和Ci分别降低了7.67%和5.02%, Ls增加14.5%, Pn下降了约19%, 说明强光和高温导致的气孔限制使净光合速率降低。而遮荫处理的光合参数变幅减小, 与自然光下生长的相比, Gs和Ci较高, Tr、tl和Ls较低, 这对叶片的光合作用非常有利。
|
|
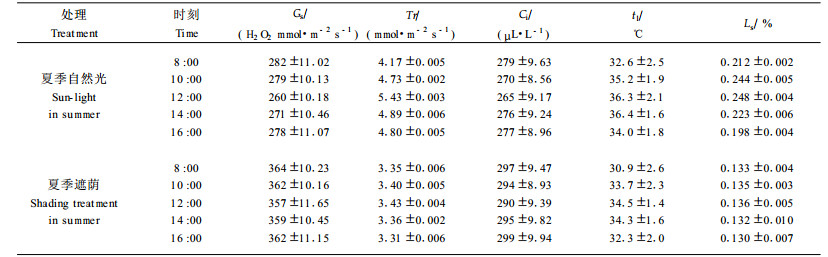
图 2显示, 夏季自然光下生长的牡丹, AQY在上午先升高后降低, 12:00左右降至最低, 而后又逐渐升高; CE则逐渐降低, 12:00左右降至最低, 下午升高。而遮荫处理后牡丹叶片的AQY和CE均比自然光下高, 中午降低不明显。LSD检验(表 2)表明, 夏季自然光与遮荫处理AQY和CE日变化在P < 0.01水平上差异极显著。说明牡丹在夏季自然光下, 强光和高温的作用使叶片的光化学效率和光合碳同化的酶活性也受到了较大的限制, 这种限制是由叶肉因素造成的, 即非气孔限制。

|
图 2 大田牡丹叶片表观量子效率(AQY)和羧化效率(CE)的日变化 Fig. 2 Diurnal variation of apparent quantum yield of photosynthesis (AQY) and carboxylation efficiency (CE) in the leaves of field Tree Peony |
|
|
AQY和Fv/Fm降低是光合作用光抑制发生的明显特征。结合图 3可见, 在夏季自然光下, Fv/Fm与Fm日变化趋势相似, 呈明显的倒“单峰”曲线, 在13:00左右降至最低, 说明自然光下, 夏季牡丹叶片的光合作用出现了明显的光抑制现象。13:00以后Fv/Fm和Fm慢慢地恢复, 16:00基本恢复到8:00水平, 说明牡丹叶片光合作用反应中心并没有被破坏, 只是反应中心PSⅡ为适应强光和高温胁迫而出现短时间的活性下调。而经夏季遮荫处理后, Fv/Fm与Fm在一天中变化幅度均明显变小。遮荫与自然光下相比, Fo没有明显的区别, 尽管在中午稍有增加, 但很快又恢复到原来的水平, LSD检验(表 2)表明, 夏季自然光与遮荫处理Fv/Fm日变化在P < 0.01水平上差异极显著。说明遮荫可明显地减轻光合作用的光抑制, 保护反应中心PSⅡ光化学反应的高水平运转。

|
图 3 大田牡丹叶片PSⅡ最大光化学效率(Fv/Fm)、初始荧光(Fo)与最大荧光(Fm)的日变化 Fig. 3 Diurnal variation of maximal PSII efficiency (Fv/Fm)、initial fluorescence (Fo) and maximal fluorescence (Fm)in the leaves of field Tree Peony |
在大田栽培条件下, 山东菏泽和河南洛阳牡丹产地4—8月的生态环境, 主要受温度和光照的影响。牡丹有庞大的肉质根系, 具有很强的吸收水分和养分的功能, 因而耐旱。繁茂的群体、根系强吸水能力促进蒸腾作用, 使空气湿度随温度、光强变化但变幅相对较小。本试验重点考虑温度、光照2个影响光合作用的主导生态因子, 以春季适宜生长的季节为对照, 研究夏季高温、强光下大田牡丹光合性能的变化趋势, 以此探讨夏季遮荫促进大田牡丹生长的光合机制。在自然条件下, 环境条件如光照和温度等都发生着规律性的变化, 植物的生命活动也随着发生变化, 叶片的Pn和AQY, 都会发生明显的日变化(Xu et al., 1997)。本试验对春季和夏季大田牡丹叶片Pn日变化的比较表明, 夏季的Pn比春季明显降低, 而且两季节Pn的日变化存在明显差异:春季和夏季遮荫处理后呈“单峰型”曲线, 中午降低较少, 而在夏季自然光下却呈“双峰型”, 中午出现明显的光合“午休”。根据Farquhar等(1982)的光合控制理论, 当Gs、Ci降低和Ls增大时, Pn的降低主要是由气孔限制引起的。在本试验中夏季自然光下生长的牡丹叶片, 当Pn出现光合“午休”时, Gs降低, Ls增加(表 1), 因此可以断定牡丹在夏季中午发生了气孔限制; 但Gs的降低(7.67%)却比Pn降低(48.98%)比例小得多, 而且CE(图 2)和Fv/Fm(图 3)在夏季中午都明显降低, 说明牡丹叶片在发生气孔限制的同时也发生了非气孔限制即叶肉限制, 与前人的研究相似(孟凡珍等, 2002;苏培玺等, 2002)。但也有不同的报道(冯建灿等, 2002), 认为CE在夏季中午降低, 是因为中午强光和高温抑制了Rubisco(RuBP羧化酶)活性从而影响了光合碳循环系统的正常运转。本研究表明, 牡丹叶片光合能力降低是强光高温的协同胁迫, 强光高温不仅影响了光合作用的光反应阶段, 也影响了光合作用的暗反应阶段。Jiang等(2001)认为, Rubisco初始活力对光合日变化影响很大, 光合“午休”涉及Rubisco初始活力的下降, 但“午休”前的光合上升和“午休”过后的光合再下降与Rubisco初始活力和Rubisco活化酶的活化与钝化有关。大田牡丹光合“午休”也可能与光合碳同化的关键酶类的日变化有关, 但尚需要进一步深入探讨。
一般认为(Demmig et al., 1987), AQY和Fv/Fm降低是光抑制最明显的2个特征。夏季自然光下生长的牡丹叶片AQY中午降至最低(图 2);Fv/Fm在中午明显降低, 下午逐渐回升, 表明PSⅡ活性下调, 出现光抑制, 但反应中心并未受到破坏, 说明强光高温下PSⅡ反应中心可逆失活。姜叶片中午也有光抑制现象, 而干旱、高温、强光等逆境条件可显著加重光抑制程度(徐坤等, 2002)。
经测定大田牡丹的饱和光强约为750 μmol·m-2 s-1, 而在夏季PAR却远远超过了牡丹的饱和光强, 这种强光不仅直接造成对光合作用的抑制, 而且还可能使叶温升高、蒸腾速率增加、大气湿度降低, 使植株水分供不应求, 从而造成气孔的部分关闭、光合作用的酶系统可逆失活。而遮荫处理后牡丹叶片Pn(图 1a)、AQY和CE(图 2)均比在自然光下高, 这可能是因为遮荫使PAR(图 1c)和ta降低(图 1d), 从而使tl降低(表 1);Gs和Ci中午降低幅度变小(表 1), 这更有利于光能利用率提高、光合作用进行和光合产物的积累; Fv/Fm和Fm在一天中的变化减小, 而Fo几乎没有变化(图 3)。这也说明是中午强光以及由强光所引起的高温使牡丹发生了光抑制。因此本研究认为, 中午Fv/Fm的降低是PSⅡ反应中心可逆失活的结果, 是牡丹对中午高温强光条件的适应性反应。在盛夏季节对牡丹植株遮荫处理, 不仅可以防止光抑制的发生或者使强光和高温所引起的光抑制减轻, 而且能够延长叶片的功能期, 一方面能够延长牡丹叶片在一天中高光合速率的持续期, 另一方面还可以使牡丹在秋季保持绿色功能期的时间延长(本试验见到, 遮荫处理能延长牡丹叶片绿色功能期15 d左右), 延缓衰老, 增加光合时间, 促进光合产物积累, 有利于牡丹壮苗培育。因此, 遮荫可以作为大田牡丹栽培的一项常规措施。
艾希珍, 张振贤, 王绍辉, 等. 2000. 强光胁迫下SOD对姜叶片光抑制破坏的防御作用. 园艺学报, 27(3): 198-201. DOI:10.3321/j.issn:0513-353X.2000.03.009 |
陈向明, 郑国生, 张圣旺. 2001. 钙对保护地栽培牡丹光合特性的影响. 园艺学报, 28(6): 572-574. DOI:10.3321/j.issn:0513-353X.2001.06.020 |
冯建灿, 张玉洁. 2002. 喜树光合速率日变化及其影响因子的研究. 林业科学, 38(4): 34-39. DOI:10.3321/j.issn:1001-7488.2002.04.006 |
付振书, 赵世杰, 孟庆伟, 等. 2005. 高温强光下耐热性不同的两个甘蓝品种幼苗光合作用差异的研究. 园艺学报, 32(1): 25-29. DOI:10.3321/j.issn:0513-353X.2005.01.006 |
郭连旺, 许大全. 1996. 田间小麦叶片光合作用的光抑制不伴随D1蛋白的净降解. 植物学报, 38(3): 196-202. |
郭晓荣, 曹坤芳, 许再富. 2004. 热带雨林不同生态习性树种幼苗光合作用和抗氧化酶对生长光环境的反应. 应用生态学报, 15(3): 377-381. DOI:10.3321/j.issn:1001-9332.2004.03.005 |
江力, 曹树青, 戴新宾, 等. 2000. 光强对烟草光合作用的影响. 中国烟草学报, 6(4): 17-20. |
蓝宝卿, 李嘉珏, 段全绪. 2002. 中国牡丹全书. 北京: 中国科学技术出版社.
|
李霞, 刘友良, 焦德茂. 2002. 不同高产水稻品种叶片的荧光参数的日变化和光适应特性的关系. 作物学报, 28(2): 145-153. DOI:10.3321/j.issn:0496-3490.2002.02.001 |
刘贤赵, 康绍忠. 2002. 不同生长阶段遮荫对番茄光合作用、干物质分配与叶N、P、K的影响. 生态学报, 22(12): 2264-2271. DOI:10.3321/j.issn:1000-0933.2002.12.034 |
孟凡珍, 张振贤, 于贤昌. 2002. 不同季节生态型大白菜光合作用对夏季高温强光的响应. 应用与环境生物学报, 8(6): 605-609. DOI:10.3321/j.issn:1006-687X.2002.06.008 |
苏培玺, 杜明武, 张立新, 等. 2002. 日光温室草莓光合特性及对CO2浓度升高的响应. 园艺学报, 29(5): 423-426. DOI:10.3321/j.issn:0513-353X.2002.05.005 |
许大全, 李德耀, 邱国雄, 等. 1987. 毛竹(Phyllostachys pubescens)叶片光合作用的气孔限制研究. 植物生理学报, 13(2): 154-160. |
徐坤, 邹琦, 郑国生. 2002. 强光下姜叶片的光呼吸及叶黄素循环. 园艺学报, 29(1): 47-51. DOI:10.3321/j.issn:0513-353X.2002.01.011 |
徐志防, 罗广华, 王爱国, 等. 1998. 强光及外源活性氧对莴苣叶绿素荧光的影响. 植物生理学报, 24(3): 279-284. DOI:10.3321/j.issn:1671-3877.1998.03.011 |
张振贤, 艾希珍, 邹琦, 等. 2000. 生姜光合效率日变化的研究. 园艺学报, 27(2): 107-111. DOI:10.3321/j.issn:0513-353X.2000.02.006 |
Barber J. 1995. Molecular basis of the vulnerability of photosystem Ⅱ to damage by light. Aust J Plant Physiol, 22: 201-208. |
Berry J A, Downton W J S. 1982. Environmental regulation of photosynthesis//Govindjee(ed). Photosynthesis: VolⅡ. New York: Academia Press, 263-343
|
Critchley C, Russell A W. 1994. Photoinhibition of photosynthesis in vivo: the role of protein turnover in photosystem Ⅱ. Plant Physiol, 92: 188-196. DOI:10.1111/j.1399-3054.1994.tb06670.x |
Demmig B, Bjorkman O. 1987. Comparison of the effect of excessive light on chlorophyll fluorescence(77k) and photon yield of O2 evolution in leaves of higher plants. Planta, 171: 171. DOI:10.1007/BF00391092 |
Farquhar G D, Sharkey T D. 1982. Stomatal conductance and photosynthesis. Ann Rev Plant Physiol, 33: 317-345. DOI:10.1146/annurev.pp.33.060182.001533 |
Jiang De'an, Lu Qing, Weng Xiaoyan, et al. 2001. Role of key enzymes for photosynthesis in the diurnal changes of photosynthetic rate in rice. Acta Agronomica Sinica, 27(3): 301-307. |
Xu Daquan, Shen Yungang. 1997. Diurnal variation in the photosynthetic efficiency in plants. Acta Phytophysiol Sin, 23(4): 410-415. |