 2006, Vol. 42
2006, Vol. 42文章信息
- 杨敏生, 李艳华, 梁海永, 王进茂.
- Yang Minsheng, Li Yanhua, Liang Haiyong, Wang Jinmao.
- 白杨派杂种无性系及其亲本光合和生长对盐胁迫的反应
- Comparison of Growth and Photosynthesis in Response to Salt Stress in Populus Hybridization Clones and Their Parent Species
- 林业科学, 2006, 42(4): 19-26.
- Scientia Silvae Sinicae, 2006, 42(4): 19-26.
-
文章历史
- 收稿日期:2004-08-03
-
作者相关文章


2. 河北省环境科学研究院 石家庄 050051
2. Hebei Provincial Academy of Environmental Sciences Shijiazhuang 050051
利用种间和种内耐盐性变异选育耐盐新品种, 是抗盐育种的有效途径。毛白杨(Populus tomentosa)是我国华北和西北平原地区主要造林树种之一, 土壤盐渍化和干旱等限制了其分布范围。将毛白杨与西北地区耐盐、抗旱白杨派树种进行有性杂交, 选育出抗盐、速生、适应性广的优良无性系, 对于充分发挥毛白杨生产潜力, 扩大其栽种范围具有重要意义。树木耐盐性的研究, 涉及到树木在高盐分逆境下的各种生理过程及外在形态表现, 乃至个体基因型的改变等, 目前已有许多相关报道(马焕成等, 1996;Morabito et al., 1996; Fung et al., 1998;Chen et al., 2002;张立钦等, 2000;张川红等, 2002), 但以抗盐品种选育为目的, 对杂种无性系及其亲本进行耐盐性比较研究还较少。实践表明:通过室内苗木耐盐生理研究, 深入探索耐盐机制; 野外定位观察, 积累经验定性判断, 两方面的有机结合对于比较和筛选抗盐品种是切实可行的。本文以速生白杨派双交杂种无性系为材料, 比较了杂种无性系与其亲本在盐胁迫条件下生理和生长的差异, 并试图找出其抗盐性的遗传规律, 为白杨派抗盐品种选育提供理论依据和方法。
1 材料与方法 1.1 植物材料选用毛白杨、新疆杨(P.alba var. pyramidalis)、毛新杨、B430共4个无性系为主要试验材料。其中毛新杨为毛白杨×新疆杨(P. tomentosa×P.alba var. pyramidalis)单交无性系, B430为双交无性系, 杂交组合为(毛白杨×新疆杨)×(银白杨×欧洲山杨)[(P. tomentosa×P.alba var. pyramidalis) ×(P. alba×P. tremula)]。
1.2 盐胁迫处理方法参试的4个无性系采用盆栽法。1999年3月下旬将4个无性系1年生嫩枝扦插苗栽于塑料盆中, 盆高30 cm, 内径25 cm, 培养土为苗圃熟土(2份)和沙(1份)混合而成, 为砂壤土。将盆下部2/3埋入土中, 在充足供水条件下培养。成活后每盆选留3株大小一致的苗木, 7月上旬进入旺盛生长期时, 开始试验。进行如下盐处理:NaCl质量浓度分别为1.0、3.0、6.0、9.0 g·L-1, 以不加NaCl的自来水为对照。每处理5盆, 试验开始后用上述不同浓度盐水处理苗木, 每隔10 d浇1次, 每次每盆浇2 L, 盐处理28 d。为防止雨水淋湿, 设置塑料棚。
1.3 指标测定 1.3.1 生长指标测定分别在盐处理后每隔7 d测定已标记苗木的苗高, 每处理测定3株, 取其平均值。施盐后, 每盆选1株苗木, 取其中正在展开的一片叶, 每隔2 d测定其叶面积。在试验结束时, 用水轻轻地冲洗土壤, 使根得以完整地洗出。选有代表性的植株, 60 ℃烘干12 h后分别测定根、茎和叶的生物量。
1.3.2 叶绿素含量测定盐处理第25天采用丙酮提取法, 用分光光度计测定各无性系叶绿素含量。
1.3.3 叶绿素a活体荧光测定NaCl处理第20天时用脉冲调制荧光仪(modulated chlorophyll fluorometer, OSL-FL, OPTISOI, USA)测定不同NaCl处理下各无性系同位叶片的荧光诱导动力学曲线。所测定叶片均为从顶部数第8片叶(为功能叶)。测前先将叶片暗适应5 min, 于上午10:00左右测定。
1.3.4 净光合速率测定盐处理第5、10、20天, 用美国产的CI-301PS光合作用测定仪于9:00左右测定各无性系的净光合速率、气孔阻力、蒸腾速率。盐处理第21天测定光合速率、蒸腾速率、气孔阻力及胞间二氧化碳浓度日变化, 第22天测定不同叶位净光合速率。
2 结果与分析 2.1 盐胁迫对生长性状的影响在不同盐胁迫条件下, 测定了4个无性系苗木叶片生长。从刚展开的幼叶开始至完全展开为止, 不同盐胁迫下, 苗木叶片生长模式基本一致, 均呈S型变化, 且单叶片从放叶到扩展到最大需10~16 d。随着盐浓度增加, 叶片生长速率迅速下降(图 1)。

|
图 1 盐胁迫对各无性系单叶片生长的影响 Fig. 1 Single leaf growth of different clones under different salt stress treatments |
在盐胁迫下各无性系苗高随时间的变化曲线基本一致, 均呈S型变化, 且随盐浓度增加, 生长曲线趋于平缓, 即生长量降低。到9.0 g·L-1NaCl处理时, 除B430和新疆杨苗高较处理前略有增加外, 毛白杨和毛新杨几乎未生长(图 2)。

|
图 2 盐胁迫对各无性系苗高生长的影响 Fig. 2 Seedling height growth of different clones under different salt stress treatments |
各无性系苗木受到盐胁迫时, 总生物量表现出减小的趋势。当盐处理浓度增加到3.0 g·L-1时, B430和新疆杨的总生物量未受到明显影响; 而毛新杨的总生物量缓慢减少, 亲本毛白杨总生物量急剧下降。当盐浓度提高到6.0 g·L-1时, 4个无性系的生物量均急剧下降。
在盐胁迫下, 根、茎、叶生物量的分配比例发生了明显变化。从表 1可以看出, 除毛白杨的根、茎、叶分配比例变化不明显外, 其余3个无性系受盐胁迫后, 根系生物量所占比例均增加, 只是增加的程度不同。B430增加的最多, 在3.0 g·L-1及9.0 g·L-1的盐处理下根系生物量占总生物量的比例分别为34.4%和51.2%, 而对照为25.1%;毛新杨相应的比例分别为25.68%、28.3%、18.4%, 新疆杨为31.9%、42.4%、24.9%。
|
|
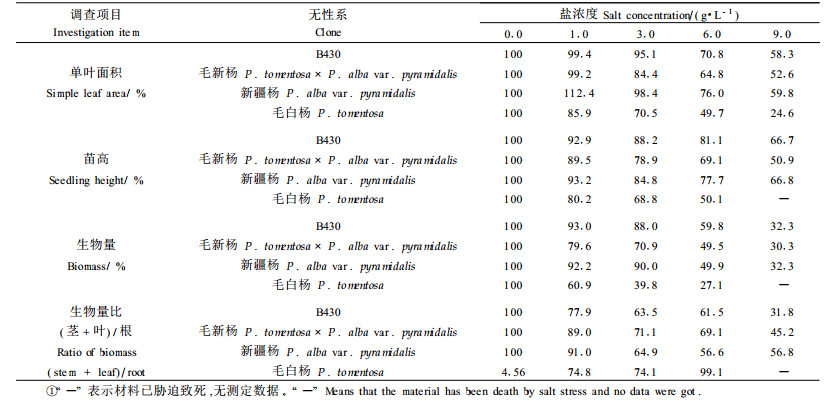
不同无性系对盐胁迫的反应不同。在盐胁迫下, 随着盐分浓度增加各生长性状均降低, 但降低程度不同。新疆杨和B430受盐胁迫影响较小, 且二者间无显著差异。而毛白杨随盐浓度增加, 苗高迅速下降。
2.2 盐胁迫对净光合速率的影响盐处理第21天测定4个无性系净光合速率(Pn)日变化, 结果(图 3)表明, 各无性系Pn具有明显的日变化规律, 4个无性系的Pn日进程基本上都呈单峰曲线, 即上午6:00—10:00时急剧上升, 至10:00时达最大值; 10:00—18:00时又迅速下降。毛白杨Pn最大值出现在8:00, 在轻度胁迫下, 一天中一直保持较高的光合能力, 但随着胁迫加强, Pn迅速下降。无性系B430、新疆杨和毛新杨, 在盐浓度为1.0~3.0 g·L-1时, 其日变化曲线下降不显著, 当大于6.0 g·L-1时日变化曲线才显著下降。如6.0 g·L-1 NaCl处理下, 10:00各无性系Pn下降程度为:毛白杨下降了66.8%, B430下降了38.3%, 新疆杨下降了30.3%, 毛新杨下降了53.5%。

|
图 3 各无性系在不同盐胁迫下叶片净光合速率日变化 Fig. 3 Diurnal variation of leaf net photosynthetic rate of different clones under different salt stress treatments |
盐胁迫处理第22天时, 各无性系净光合速率随叶顺序变化见图 4。由图 4可以看出, 整个枝条自顶部至基部不同叶位叶片的光合速率呈单峰曲线, 中部叶片最高, 两端叶片净光合速率渐低。6.0 g·L-1的盐胁迫对4个无性系嫩叶光合速率影响都不大。但是, 盐胁迫明显降低4个无性系的中部叶及老叶的净光合速率。6.0 g·L-1的NaCl处理下, 毛白杨有落叶现象, 且叶片生长受到盐离子的抑制。毛新杨和B430也有轻微的落叶, 而新疆杨基本无落叶现象。

|
图 4 盐胁迫下各无性系不同叶位净光合速率变化 Fig. 4 Net photosynthetic rate of different leaf position under different salt stress treatments |
图 5为盐胁迫处理第5天和第20天时, 4个无性系功能叶片上午9:00的净光合速率。1.0~6.0 g·L-1 NaCl处理5 d后, 4个无性系的Pn值均缓慢下降。9.0 g·L-1 NaCl处理下, Pn值迅速下降, 尤其是毛白杨Pn值急剧下降趋于0。盐处理20 d时, 4个无性系的Pn值下降速率较第5天均加快。毛白杨下降最快, 毛新杨和新疆杨下降速率居中, B430下降较缓慢。B430在9.0 g·L-1 NaCl处理下, Pn值仍高达5.8 μmol·m-2s-1。

|
图 5 不同胁迫时间对各无性系功能叶片净光合速率的影响 Fig. 5 Influence of salt stress time on leaf net photosynthetic rate of different clones |
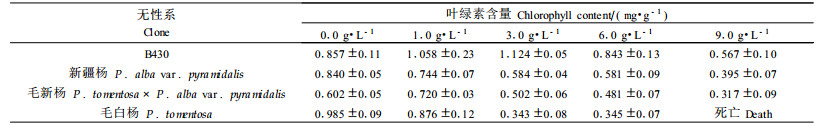
从表 2可以看到, 随着盐浓度的增加, 亲本毛白杨和新疆杨的叶绿素含量逐渐减少, 但二者下降幅度不一样, 毛白杨比新疆杨下降速度明显快。B430和毛新杨在低盐浓度下, 叶绿素含量上升, 高盐处理下才开始下降。在6.0 g·L-1的盐处理下, 4个无性系的叶绿素含量均开始下降, 此时B430叶绿素含量为对照的98%, 新疆杨为69%, 毛新杨为80%, 毛白杨仅为35%。2个杂种无性系又以B430表现较好, 在盐浓度为6.0 g·L-1时叶绿素含量才开始下降。
|
|
测定了盐胁迫20 d时各无性系在“Kintic”模式下的荧光诱导动力学曲线, 并依据此曲线算出了荧光诱导动力学参数qP(荧光光化学猝灭系数)、qN (荧光非光化学猝灭系数)及Fv/Fo(PSⅡ潜在活性)。
研究结果表明, 无性系经盐分处理后, 其荧光诱导动力学曲线发生明显变化, 随着处理浓度的增大, 曲线中的荧光诱导最高峰P相明显下降, M相(second maximum)也逐渐降低直到消失, P相的降低说明盐分引起PSⅡ反应中心活性有部分丧失, 使得稳定电子受体Q的还原受到阻碍, 因而可变荧光降低; 另外, 由于由S相(quasisteady state)向M相的上升是与O2的释放速度相平衡的(朱新广等, 1999b), 因此, M相的完全消失说明PSⅡ氧化已受到完全破坏, 表现在曲线上的这种变化说明了盐害已使叶片的结构和功能遭到部分破坏。
qP猝灭是由QA-等再氧化所造成, 是QA氧化态的一种度量, 表示PSⅡ反应中心开放部分的比例, 因而它与光合电子传递、光合放氧等过程直接相关(Van kooten et al., 1990)。在加强饱和光使荧光值达到“P”峰时, 荧光光化学猝灭系数qP值最小, 然后随时间缓慢上升, 直到处于一个稳定值, 其变化反映了PSⅡ原初电子受体QA由完全还原状态重新氧化的过程。图 6为不同盐胁迫下qP稳定值的变化。随着盐浓度的升高, 4个无性系的最大qP均逐渐下降, 但变化不大。特别是B430和毛新杨下降程度和速率均很小。在9.0 g·L-1盐分处理下, B430和毛新杨qP值较对照才分别下降了16.3%和16.7%, 新疆杨下降了22.4%, 毛白杨下降了24.6%, 这可能是因为qP比较稳定。

|
图 6 盐胁迫对各无性系qP和qN的影响 Fig. 6 Influence of salt tress on qP and qN of clones |
qN为荧光非光化学猝灭系数, 反映PSⅡ反应中心非辐射能量耗散能力的大小。qN随时间变化的动力学过程表现为先上升后下降, 直到一个较稳定的值。qN稳定时各无性系对盐胁迫的反应见图 6。由图 6可以看出, 在盐胁迫下4个无性系qN值均随盐浓度的增加而上升。新疆杨、毛新杨、B430在低盐(<3.0 g·L-1)处理时, qN值上升缓慢, >3.0 g·L-1时才迅速上升。而亲本毛白杨在低盐时即迅速上升。
从表 3可以看到, 盐胁迫降低了4个无性系叶片的Fv/Fo比值, 表明盐胁迫降低了叶片PSⅡ光化学活性, 使光系统Ⅱ受到了损伤。随着盐浓度的提高, 毛白杨Fv/Fo比值下降程度及下降速率最大, B430则相反, 下降缓慢, 毛新杨和新疆杨下降速率适中, 毛新杨略好于新疆杨。由此说明, 毛白杨光系统Ⅱ受损伤最严重, B430受损伤最轻。
|
|

在盐胁迫条件下, 植物具有最大的生产力是抗盐碱育种的根本目的, 因此, 在盐胁迫条件下, 高生长量是树木抗盐碱育种的重要选择指标。但是, 对于树木来说, 仅根据苗期的生长, 特别是盆栽苗的生长来进行选种是不可靠的。将胁迫和非胁迫条件下苗木的生长相比较, 则能较好地反映出植物对盐胁迫的适应能力, 这在农业抗盐品种选育中已得到广泛的应用。表 1结果表明, 盐胁迫下, 各无性系的单叶面积、苗高和生物量生长均受到明显影响, 特别是盐浓度达到6.0 g·L-1时, 受到明显抑制。但各无性系下降的幅度不同, B430和新疆杨下降幅度最小, 毛白杨下降幅度最大, 在盐浓度为9.0 g·L-1胁迫28 d时, 毛白杨苗木已经死亡。
在盐胁迫下根系生物量是植物适应盐胁迫的重要标志。当植物受到盐胁迫时, 根系生物量在整个植物生物量中所占比例提高, 有助于植物从土壤中吸收水分, 保持体内水分平衡。此外, 根系生物量所占比例提高, 可以使根部储存较多的Na+, 以阻止Na+在地上部积累, 从而减少Na+对叶片的伤害, 有利于增加植物的耐盐性(杨敏生等, 2003; Garcia-Sanchez et al., 2002)。有报道表明, 非盐生植物受盐胁迫后最早的反应是叶片生长变得缓慢, 与叶(包括茎)的生长相比, 根的生长总是很少或几乎不受影响, 于是根冠比上升(Chartzoulakis et al., 2002; Fung et al., 1998)。但也有研究指出, 茎和根对NaCl同样敏感, 盐胁迫导致茎和根的干质量下降的比例一样, 于是根冠比并没有变化(Seemann et al., 1985; Chartzoulakis, 1994)。在本项研究中, 盐胁迫下4个无性系苗木根、茎、叶各部分干物质积累逐渐受到抑制, 但茎和叶生物量受抑制最明显, 根生物量受影响较小, 因此随着盐胁迫的发展, 根与茎和叶的生物量之比表现为明显的上升趋势。根冠比除毛白杨无明显变化外, 其余3个无性系的根冠比均随盐浓度提高而上升。
光合同化速率降低是盐胁迫下生长受抑制的重要原因。盐胁迫降低光合速率, 减小同化物与能量供给, 从而限制植物的生长发育(Parida et al., 2005; Mansour et al., 2004;朱新广等, 1999a), 但不同树种受影响程度不同(张川红等, 2002;Morabito et al., 1996; Chartzoulakis et al., 2002)。本研究表明, 当盐浓度达到6.0 g·L-1时, 各无性系光合作用即受到明显影响, Pn开始显著下降, 且随着胁迫时间延长, 下降的幅度加大。在4个无性系中, B430的Pn对盐胁迫的反应最不明显, 而毛白杨反应最敏感, 在盐浓度达到3.0 g·L-1时即已受到明显影响。苗木受到盐胁迫后, 不同叶位Pn变化模式没有发生改变, 叶位从嫩到老, 其Pn曲线为单峰型, 即中部功能叶Pn值最大, 嫩叶和老叶较小。但盐胁迫下叶片光合能力受到显著抑制。盐胁迫对嫩叶Pn值几乎没有影响, 却显著降低老叶和中部功能叶的Pn, 这可能是因为中下部盐离子含量较高, 破坏了叶绿素, 从而降低Pn值。NaCl影响光合作用主要有3种可能的途径, 即离子伤害、渗透伤害和糖分积累造成反馈抑制, 但盐胁迫影响光合作用的根本原因, 目前还没有形成统一的认识(朱新广等, 1999a)。
植物体内叶绿素荧光诱导动力学参数对外界各种胁迫因子均十分敏感, 因而有可能将它作为植物理想的各种抗逆性指标(张守仁, 1999;史胜青等, 2004)。盐胁迫降低各无性系qP值, 说明盐胁迫降低了PSⅡ反应中心开放部分的比例, 即增加了PSⅡ关闭部分的比例, 从而抑制叶片PSⅡ光合电子传递能力, 限制了为光合碳同化提供更充足的ATP和NADPH, 这可能是盐胁迫下Pn下降的一个原因。在盐胁迫下, 4个无性系qN值均表现为上升趋势, 这表明PSII的潜在热耗散能力增强, 通过非辐射型热耗散消耗光捕获蛋白复合物(LHCII和LHCI), 吸收过剩光能而避免对光合器官的损伤(Barder et al., 2000)。这一研究结果与其他报道相似, 朱新广等(1999b)研究表明, 在较低的光强下, NaCl处理的冬小麦(Triticum aestivum), 其荧光光化学猝灭效率qP较低, 荧光非光化学猝灭效率qN较高, Fo猝灭系数较大, QB-非还原性PSII反应中心含量较大; 而在较高光强下, 其荧光非光化学猝灭效率和Fo猝灭系数则相对较低。
收集、鉴定抗盐亲本材料, 采用杂交育种手段, 促进有利基因的重组, 是培育抗盐新品种的有效途径。杂种无性系间耐盐能力差异, 与它们之间的亲缘关系有关。毛新杨是毛白杨与新疆杨的杂种, B430为毛新杨的双交杂种, 其杂交组合是(毛白杨×新疆杨)×(银白杨×欧洲山杨)。新疆杨和银白杨是我国西北地区主要杨树造林树种, 耐盐性较强, 而毛白杨主要分布在华北和西北东部, 耐盐能力较差。上述研究结果与实际情况相符, 而杂种B430则继承了新疆杨和银白杨的耐盐特性, 其耐盐性与亲本新疆杨一致, 另一单交杂种毛新杨耐盐性则介于亲本新疆杨和毛白杨之间。
马焕成, 冯衍枝, 王沙生, 等. 1996. 胡杨抗盐机理初探. 中国林学, 15(2): 31-40. |
史胜青, 袁玉欣, 杨敏生, 等. 2004. 水分胁迫对4种苗木叶绿素荧光的光化学猝灭和非光化学猝灭的影响. 林业科学, 40(1): 168-173. DOI:10.3321/j.issn:1001-7488.2004.01.028 |
杨敏生, 李艳华, 梁海永, 等. 2003. 盐胁迫下白杨无性系苗木体内离子分配及比较研究. 生态学报, 23(2): 271-278. DOI:10.3321/j.issn:1000-0933.2003.02.008 |
张立钦, 郑勇平, 吴纪良, 等. 2000. 黑杨派新无性系水培苗对盐胁迫反应的研究. 浙江林学院学报, 17(2): 121-125. DOI:10.3969/j.issn.2095-0756.2000.02.001 |
张守仁. 1999. 叶绿素荧光动力学参数的意义及讨论. 植物学通报, 16(4): 444-448. DOI:10.3969/j.issn.1674-3466.1999.04.021 |
张川红, 沈应柏, 尹伟伦, 等. 2002. 盐胁迫对几种苗木生长及光合作用的影响. 林业科学, 38(2): 27-31. DOI:10.3321/j.issn:1001-7488.2002.02.006 |
朱新广, 张其德. 1999a. NaCl对光合作用影响的研究进展. 植物学通报, 16(4): 332-338. |
朱新广, 张其德, 匡廷云. 1999b. NaCl胁迫对PSII光能利用和耗散的影响. 生物物理学报, 15(4): 780-790. |
Barder M R, Ruuska S, Nakano H. 2000. Electron flow to oxygen in higher plants and algae: rates and control of direct photoreduction (Mehler reaction) and rubisco oxygenase. Biological Sciences, 1402: 1433-1445. |
Chartzoulakis K, Loupassaki M, Bertaki M, et al. 2002. Effects of NaCl salinity on growth, ion content and CO2 assimilation rate of six olive cultivars. Scientia Horticulturae, 96: 235-247. DOI:10.1016/S0304-4238(02)00067-5 |
Chartzoulakis K S. 1994. Photosynthesis, water relations and leaf growth of cucumber exposed to salt stress. Scientia Horticulturae, 59: 27-35. DOI:10.1016/0304-4238(94)90088-4 |
Chen S, Li J, Fritz E, et al. 2002. Sodium and chloride distribution in roots and transport in three poplar genotypes under increasing NaCl stress. Forest Ecology and Management, 168: 217-230. DOI:10.1016/S0378-1127(01)00743-5 |
Fung L E, Wang S S, Altman A, et al. 1998. Effect of NaCl on growth, photosynthesis, ion and water relations of four poplar genotypes. Forest Ecology and Management, 107: 135-146. DOI:10.1016/S0378-1127(97)00328-9 |
Garcia-Sanchez F, Jifon J L, Carvajal M. 2002. Gas exchange, chlorophyll and nutrient contents in relation to Na+and Cl- accumulation in 'Sunburst' mandarin grafted on different rootstocks. Plant Science, 162: 705-712. DOI:10.1016/S0168-9452(02)00010-9 |
Morabito D, Jolivet Y, Prat D, et al. 1996. Differences in the physiological responses of two clones of Eucalyptus microtheca selected for their salt tolerance. Plant Science, 114(2): 129-139. DOI:10.1016/0168-9452(96)04325-7 |
Mansour M M F, Salama K H A. 2004. Cellular basis of salinity tolerance in plants. Environmental and Experimental Botany, 52: 113-122. DOI:10.1016/j.envexpbot.2004.01.009 |
Parida A K, Das A B. 2005. Salt tolerance and salinity effects on plants: a review. Ecotoxicology and Environmental Safety, 60: 324-349. DOI:10.1016/j.ecoenv.2004.06.010 |
Seemann J R, Critchley C. 1985. Effects of salt stress on the growth, ion content, stomatal behaviour and photosynthetic capacity of salt-sensitive species Phaseohs vulgaris L. Planta, 164: 151-162. DOI:10.1007/BF00396077 |
Van kooten O, Snel J F H. 1990. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosyn Res, 25: 147-150. DOI:10.1007/BF00033156 |