 2006, Vol. 42
2006, Vol. 42文章信息
- 杨锦昌, 许煌灿, 尹光天, 曾炳山, 冯昌林.
- Yang Jinchang, Xu Huangcan, Yin Guangtian, Zeng Bingshan, Feng Changlin.
- 单叶省藤组培家系的生长特性分析
- An Analysis on Growth Character of Tissue Culture Families of Calamus simplicifolius
- 林业科学, 2006, 42(2): 120-124.
- Scientia Silvae Sinicae, 2006, 42(2): 120-124.
-
文章历史
- 收稿日期:2003-06-18
-
作者相关文章



2. 中国林业科学研究院热带林业 实验中心 凭祥 532600
2. Experimental Center of Tropical Forestry, CAF Pingxiang 532600
我国制藤工业发达、工艺精良,藤产品出口约占国际市场份额20%。然而国内棕榈藤资源匮乏,极大制约了制藤工业和出口贸易的发展。近年来,为了摆脱资源危机的被动局面,加快了棕榈藤资源人工培育的步伐(许煌灿等,2001)。单叶省藤(Calamus simplicifolius)是中国特有棕榈藤种之一,藤茎具良好工艺特性,是藤编家具及工艺品的优良材料,是华南地区推广栽培的首选藤种(江泽慧,2002;曾炳山等,2003)。良种先行是发展棕榈藤人工林的首要步骤,而通过组培快繁苗木则是良种先行的重要途径。然而与其他林木相比,国内外对棕榈藤的研究较迟(许煌灿等,1994;Dransfield,2002),迄今为止还没有关于棕榈藤家系选优的研究报道。最近,国内学者对单叶省藤组培苗造林开始了初步研究,为良种选育提供了一定的借鉴(曾炳山等,2003)。然而该研究材料来源于幼龄林,对于组培苗造林后期的生长性状缺乏必要的分析。因此,利用长期观测的固定标准地材料,对单叶省藤组培家系的生长特性进行分析,从而为组培家系的选优提供依据,是一项十分有意义的工作。
1 材料与方法 1.1 试验地概况试验地位于广西凭祥市热带林业实验中心,北纬22°07′27″,东经106°44′30″,海拔210 m,属南亚热带季风气候,干湿季节交替明显。年平均气温21.4 ℃,最冷月份平均气温13 ℃,极端最低气温-1.5 ℃,最热月份平均气温27.5 ℃,极端最高气温39.8 ℃,≥10 ℃积温7 818.4 ℃,年平均降水量1 379 mm,蒸发量1 300~1 700 mm。试验地地势平坦开阔,土壤肥力比较均匀,发育母岩为石灰岩,土层厚度>1 m,pH值6.4,有机质含量9.8 g·kg-1,全N 0.7 g ·kg-1,速效N为60.4 mg·kg-1,全P 0.42 g·kg-1,速效P 6.0 0 mg·kg-1,全k 5.4 g·kg-1,速效K 55.8 mg·kg-1。
1.2 试验设计1) 造林措施 造林前,全面清理林下杂草和灌木,适当疏伐林木,控制上层林分郁闭度约0.5,穴状整地,种植穴规格为50 cm×50 cm×30 cm,每穴种植1株藤苗;造林苗木为1年生藤苗。2)试验安排 在热带林业实验中心树木园引种区设置组培家系和实生苗试验。采用完全随机区组设计安排4个组培家系(即CS10、CS11、CS3和CS5)和1个实生苗对照(CK),2次重复,每区组5个试验小区。试验小区为长方形,每小区种植20株,株行距3 m×2 m。每小区两侧设置保护行。3)数据采集和处理 造林前测定各家系的苗高和叶片数;造林后2.5、4、5、和9年时测定保存率、萌蘖数和长度。数据采集后,对保存率和萌蘖率与对茎长分别进行sin-1x1/2转换和lnx转换,并在计算机上利用SPSS统计软件进行处理和分析。
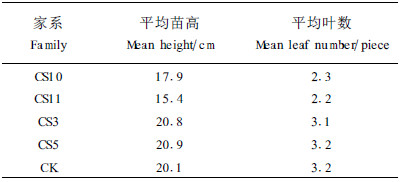
2 结果与分析 2.1 造林前藤苗生长性状不同家系的遗传成分不尽相同,会影响到苗期的生长表现。从表 1可知,在4个组培家系中,CS3和CS5表现最好,CS10次之,而CS11表现最差。方差分析结果表明:不同家系间苗高达到极显著差异(见表 2)。经多重比较分析,CS10、CS11与CS3、CS5之间存在极显著差异;CS10与实生苗也存在显著差异,而CS11与实生苗达到极显著差异。该结果说明:不同组培家系的苗高在幼龄时存在极显著差异,苗期生长性状在家系间存在较大的变异。
|
|
|
|

保存率是衡量造林成败的关键指标,同时也是体现植株在整个生长过程中对外界环境适应能力。由图 1可知,各家系保存率随着年龄的增加开始时下降比较迅速,接着变化速度减缓,最后处于稳定状态。根据这一特征,可将植株生长大致分为3个阶段:A适应阶段(苗龄1~4年):这段时间内,各个家系保存率的变化幅度即折线的斜率均比较大;其中CS11保存率最高,在2.5年和4年时分别达到97.5%和90%;CS5和实生苗保存率在4年时最低,保存率均小于80%;而CS10和CS3的保存率较高。B过渡阶段(4~5年):除CS3之外,组培家系保存率的下降逐渐减缓甚至不变,而实生苗的保存率则保持不变;此时CS11的保存率虽然最高,但已下降至82.5%;而实生苗最低,仅为75%。C稳定阶段(苗龄≥5年):各家系的保存率不再下降。总之,各组培家系的保存率存在一定的差异,但到后期它们的保存率均高于实生苗,经方差分析表明,各家系的植株保存率均无显著差异。

|
图 1 各家系保存率随年龄变化规律 Fig. 1 Changes in survival rate of each family with ages |
萌蘖是棕榈藤的重要生物学特性和重要生长指标,它促使植株不断分蘖、导致植株数量不断增加、进而增加藤丛的总茎长(曾炳山等,2003;许煌灿等,1994)。萌蘖特性可体现在萌蘖起始时间、萌蘖率和平均萌蘖数3个方面。1)萌蘖起始时间即造林后植株开始萌蘖的时间。从表 3可知,组培家系的萌蘖起始时间比较一致,除CS3在4年萌蘖稍迟外,其他3个家系均在2.5年开始萌蘖。而实生苗萌蘖起始时间为5年;相比之下,4个组培家系的萌蘖起始时间均早于实生苗1~2年, 这表明用组培苗造林更有利于植株提早萌蘖。2)萌蘖率和平均萌蘖数萌蘖率是指已萌蘖的藤丛数与所有藤丛数的百分比,而平均萌蘖数是指植株已分蘖的植株数量与所有藤丛数的比值,二者联系紧密,但又有区别。前者侧重于描述植株是否萌蘖的一种状态,体现植株萌蘖的齐性;后者则反映植株萌蘖的速度,是对前者的进一步补充和丰富。
|
|
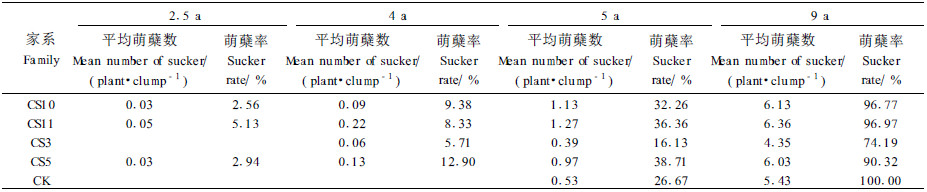
当植株年龄为2.5年时,除CS3未出现萌蘖外、其他3个组培家系均已开始萌蘖,但萌蘖率和平均萌蘖数都很低。当年龄为4年时,除实生苗外,所有组培家系全部开始萌蘖,但萌蘖率和萌蘖数量仍很有限,只有CS5的萌蘖率较高,达到12.9%;CS11的萌蘖数较多,平均为0. 22个·丛-1;而其他两个家系的萌蘖率和平均萌蘖数都很低。随着时间推移至5年时,所有家系都已开始萌蘖。此时,CS10、CS11和CS5家系的萌蘖速度不断加快,平均萌蘖数和萌蘖率均高于实生苗,分别增长到32%和0.9个·丛-1以上,这表明近1/3以上的藤丛产生分蘖,而且每个藤丛萌蘖1株以上;但CS3的萌蘖性最差,其萌蘖率和平均萌蘖数均小于实生苗。到了9年时,各家系进一步加快萌蘖生长,除CS3外,其他3个组培家系的平均萌蘖数和萌蘖率分别在6个和90%以上,虽然实生苗家系萌蘖率达100%,但平均萌蘖数较之于CS11、CS10和CS5低。从总体上看,4个组培家系中CS11的萌蘖性最强,CS5和CS10次之,CS3最弱。
2.4 株数变化萌蘖力的差异直接影响到各家系藤丛株数的变化。通过绘制各家系总株数变化图(图 2)可知,在1~4年阶段,除CS11的株数略有增长之外,其他4个家系的株数均比造林时的株数有所下降,这是因为大部分的家系刚刚萌蘖或还未萌蘖,由萌蘖所增长的植株数小于造林后死亡数量。在4~5年时期,各家系的藤株数均大于造林时的株数,这主要归结于萌蘖速度的加快和造林保存率的逐渐稳定,但各家系间有所差异:CS11藤株数的增长仍然最大,CS5和CS10植株数目增长较多,而CS3和实生苗家系在植株数量上增长最慢。随着年龄增至9年时,各家系的株数迅速增多,但各家系植株数量增长的差异进一步增大:CS11依旧维持最高水平,CS10和CS5二者几乎并列处于第2,实生苗第3,而CS3最低。方差分析结果表明,各家系间株数差异不显著。

|
图 2 各家系株数随年龄的变化规律 Fig. 2 Changes in tree number of each family with ages |
随着年龄的增加,藤丛不仅横向萌蘖扩展,而且纵向攀援伸长,导致藤丛茎长不断发生变化。藤丛茎长涉及母茎长、萌蘖茎长和总茎长,后者是前两者的相加结果。由于不同家系的萌蘖力的差异,其萌蘖茎长的变化规律不太容易把握,因而本文将着重讨论母茎长和总茎长的变化规律。从图 3、4可见,各家系在年龄≤5年时平均母茎长和藤丛总茎长均很小,二者的数值皆小于1 m,而且增长非常缓慢,3个点的连线几乎是一条平直的直线。而后随年龄增加,点的连线明显变陡,平均母茎长和平均藤丛总茎长增长迅速;同时,各家系的增长趋势逐步明显,形成2类折线簇,即由CS10、CS11和实生苗组成的第1类折线簇和由CS3和CS5组成的第2类折线簇。这种现象表明随时间后移,各家系在茎长指标上的差异进一步增大,并分化形成两大类,这为单叶省藤的家系选优提供了参考和依据。

|
图 3 各家系平均母茎长变化规律 Fig. 3 Change in mean length of mother stem of each family |
对藤丛母茎长和藤丛总茎长进行方差分析(见表 4)可知:在不同年龄时,母茎长在家系间均达到极显著差异,总茎长在家系间除年龄为5年时只达到显著差异水平外,其他年龄段均达到极显著差异,这表明进行单叶省藤家系选择试验研究是十分必要的。

|
图 4 各家系藤丛总茎长变化规律 Fig. 4 Change in mean total length of rattan clump of each family |
|
|
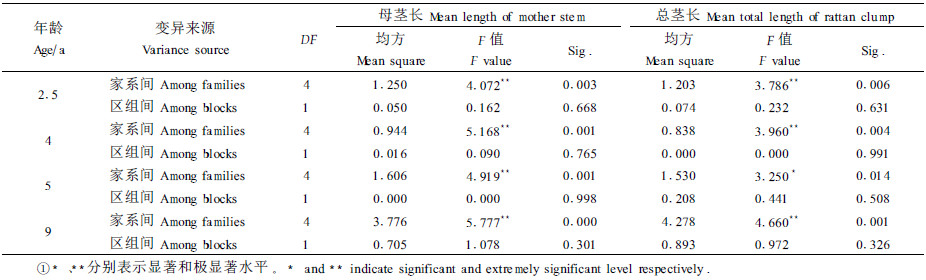
采用S检验法对年龄为9年时的平均母茎长和藤丛总茎长进行多重比较,结果得知:在平均母茎长指标上,CS5、CK与CS3、CS10之间以及CS11与CS10之间均存在极显著差异;在藤丛总茎长指标上,CS5、CK与CS3、CS10之间存在极显著差异,而CS11与CS3、CS10之间存在显著差异。
3 结论与讨论本次试验的4个组培家系材料均来自同一棕榈藤林分,但通过苗期生长性状分析,不同组培家系在苗高指标上存在极显著差异,表明组培子代在苗期生长性状上会出现较大的变异,藤类的无性系和家系选育可望获得较大的遗传增益(曾炳山等,2003)。藤苗的叶片数因不满足条件而未进行方差分析,但通过相关关系分析,得出藤苗的叶片数与苗高呈极显著正相关,Spearman相关系数达到0.530,说明叶片数可以作为衡量苗木生长表现的指标。另外,棕榈藤的苗木与其他树种不同(杨宗武等,2003;何贵平等,2003),藤苗在幼龄时期地径不规则而在测量时容易出现较大误差,因而在苗期生长性状分析中未采用地径指标。
在植株生长过程中,各家系的保存率在年龄≤4年下降迅速,而后变化幅度逐步减小并最后处于稳定状态,表明藤苗刚造林后由于苗木小,对不良因素的抗逆性和对外界环境的适应性较差,而后随年龄增长,植株的抗逆性和适应性不断增强,最终使得保存率维持在一个稳定水平;另一方面,可以反映出:藤苗造林后头3年要加强抚育管理工作,为苗木创造一个良好的生长环境,促使藤幼龄林尽早郁闭(江泽慧,2002;许煌灿等,1994;Nur Supardi et al., 1992)。家系间的保存率虽有一定的差异,但无论是区组间还是家系间,均未达到显著水平, 一方面说明试验地的林分透光度、立地条件和造林措施比较一致;另一方面可能与组培的母株均来自同一人工林有关。从整个过程看,家系的保存率均在75%以上,说明采用营养袋培育的藤苗比较强壮,可以较好地承受大田移栽并保持较高的成活率。
组培家系的萌蘖起始时间均早于实生苗,时间也比较一致,而且除CS3之外,其他3个组培家系的平均萌蘖数和平均萌蘖率皆大于实生苗。可以认为组培家系的萌蘖力强于实生苗, 这种现象可从两个方面加以解释:其一,组培苗在继代培养时激素不断积累,这些激素累积所产生效应会影响到组培苗造林后的分蘖(苏秀城,2000);其二,组培家系所选择的母株也是影响组培子代的因素,因为通过无性繁殖,组培子代的遗传成分与其原株完全相同(梁一池,1997)。组培家系萌蘖力影响到组培家系的总株数。从前面的分析可知各家系的萌蘖特性不论是从平均萌蘖数还是萌蘖率,基本上都是CS11>CS10>CS5>CK>CS3。
藤丛母茎长和藤丛总茎长这两个指标在家系间的显著差异或极显著差异反映出组培家系间的变异程度。藤丛母茎长与总茎长的生长规律并不完全一致,平均母茎长最长的并不一定反映平均总茎长最长;反之亦然。这可归结于家系萌蘖特性的差别,因为萌蘖力牵涉到藤丛株数变化,进而影响到藤丛的总茎长。根据方差分析和在9年时的多重比较结果,可最终将4个组培家系归为2类,即一类是由CS5和CS11优良组培家系与CS3和CS10组成的普通组培家系。
苗期时组培家系的生长表现与造林后期的生长表现会产生一些差异,如CS3在苗期时生长表现很好,但到了9年时则表现最差;而CS11苗期表现最差,但在后期却生长迅速。由此可见,苗期生长性状并不完全决定造林后期植株的生长表现(余荣卓, 1998;李明鹤等,2003)。由于林木是多年生的植物,林木的种源家系试验往往得经过连续数年或更长时间才能较明显地得出结果,这样也有利于做出更为客观和准确的分析与判断(洪伟,1997;赵承开,2002)。组培家系无论是在苗期还是在造林后的生长差异表明了对不同家系进行生长性状比较和分析是非常有必要的,也表明了设置家系试验进行家系选优的合理性。
本文所分析的组培家系仅限有4个,这与家系选优时所需要的数量相差较远。由于我国棕榈组培研究开始于20世纪90年代(庄承纪等,1991;张方秋,1993;许煌灿等,2002),许多技术尚处于摸索阶段,再加上条件限制,而未能多地点、多样本选择组培母株加大组培家系数量。今后应该增加组培家系的数量,拓宽和丰富优良基因资源,开展种源和家系综合选择的研究(徐建民等,2003),使得研究结果更有代表性和科学性。
何贵平, 陈益泰, 孙银祥, 等. 2003. 南酸枣地理种源期性状变异研究. 林业科学研究, 16(2): 177-182. DOI:10.3321/j.issn:1001-1498.2003.02.010 |
洪伟编著.1997.林业试验设计技术与方法.北京: 科学技术出版社, 205-211
|
江泽慧主编.2002.世界竹藤.沈阳: 辽宁科学技术出版社, 592-593
|
李明鹤, 沈宝仙, 李传涵, 等. 2003. 杉木无性系选择与木材生产相结合的研究. 林业科学, 39(3): 157-162. |
梁一池. 1997. 林木育种原理与方法. 厦门: 厦门大学出版社, 187-195.
|
苏秀城. 2000. 杉木无性不同继代代数组培差异研究. 福建林学院学报, 20(4): 353-356. DOI:10.3969/j.issn.1001-389X.2000.04.018 |
徐建民, 陆钊华, 李光友, 等. 2003. 细叶桉种源—家系综合选择的研究. 林业科学研究, 16(1): 1-7. DOI:10.3321/j.issn:1001-1498.2003.01.001 |
许煌灿, 孙清鹏.2001.棕榈藤产业现状及其发展前景//竺肇华主编.中国热带地区竹藤发展.北京: 中国林业出版社, 88-106
|
许煌灿, 尹光天, 孙清鹏, 等. 2002. 棕榈藤的研究和发展. 林业科学, 38(2): 135-143. DOI:10.3321/j.issn:1001-7488.2002.02.023 |
许煌灿, 尹光天, 曾炳山主编.1994.棕榈藤的研究.广州: 广东科技出版社
|
杨宗武, 郑仁华, 侯伯鑫, 等. 2003. 福建柏苗期生物量种源间遗传变异及综合评价的研究. 林业科学研究, 16(1): 39-44. DOI:10.3321/j.issn:1001-1498.2003.01.007 |
余荣卓. 1998. 杉木种子园子代长期测定结果与早晚期选择相关性分析. 福建林学院学报, 18(4): 362-365. DOI:10.3969/j.issn.1001-389X.1998.04.020 |
曾炳山, 尹光天, 许煌灿, 等. 2003. 单叶省藤组培苗造林初步研究. 林业科学研究, 16(2): 240-244. DOI:10.3321/j.issn:1001-1498.2003.02.019 |
张方秋. 1993. 棕榈藤组培技术研究. 林业科学研究, 6(5): 486-492. |
赵承开. 2002. 杉木优良无性系早期选择年龄和增益. 林业科学, 38(4): 53-60. DOI:10.3321/j.issn:1001-7488.2002.04.009 |
庄承纪, 周建葵. 1991. 省藤组织培养的植株再生. 云南植物研究, 13(1): 97-100. |
Drandfield J. 2002. General introduction to rattan—the biological background to exploitation and the history of rattan research//Dransfield John, Florentino O Tesoro and Manokaran N (ed). Rattan current research issues and prospects for conservation and sustainable development.FAO, 23-34
|
Nur Supardi M N and Aminuddin M. 1992. Planting systems for large_diameter canes//Razali W, Dransfield J, Manokaran N(ed). A guide to the cultivation of rattans.Malaysian Forest Records No. 35. Forest Research Institute Malaysia, 89-98
|