 2006, Vol. 42
2006, Vol. 42文章信息
- 李新彬, 王襄平, 罗菊春, 牟长城, 林英建, 夏新莉.
- Li Xinbin, Wang Xiangping, Luo Juchun, Mu Changcheng, Lin Yingjian, Xia Xinli.
- 干扰对长白山北坡次生林群落结构和树种多样性的影响
- Influence of Human Disturbance on Community Structure and Tree Species Diversity of Secondary Forests on Northern Slope of Mt. Changbai
- 林业科学, 2006, 42(2): 105-110.
- Scientia Silvae Sinicae, 2006, 42(2): 105-110.
-
文章历史
- 收稿日期:2004-12-21
-
作者相关文章



2. 北京大学环境学院生态学系 北京 100871;
3. 东北林业大学林学院 哈尔滨 150040;
4. 吉林省白河林业局黄松蒲林场 安图 133613;
5. 北京林业大学生物科学与技术学院 北京 100083
2. Department of Ecology, College of Environmental Sciences, Peking University Beijing 100871;
3. College of Forestry, Northeast Forestry University\Harbin 150040;
4. Huangsongpu Forest Farm, Baihe Forestry Administration of Jilin Province Antu 133613;
5. College of Biological Sciences and Biotechnology, Beijing Forestry University Beijing 100083
阔叶红松(Pinus koraiensis)林为中国东北东部山地物种最丰富、生产力和生物量最高的森林群落类型,具有结构复杂、土壤肥力高、群落稳定性强的特点(徐振邦, 1985)。长白山区是阔叶红松林的核心分布区(吴方正, 2003),但由于多年的采伐和其他人为干扰,形成了大面积的次生林。
对阔叶红松林的群落结构和植物多样性已有大量研究(王战等, 1980;徐化成, 2001;郝占庆,2000),在自然干扰对阔叶红松林的影响方面,也有一定的研究(阳含熙等, 1994;臧润国,1995),但关于次生林的研究相对较少。有研究认为次生林物种多样性低于阔叶红松林(雷相东等, 2003),也有研究发现干扰较小的次生林多样性更高(郝占庆等, 1994;罗菊春等, 1997;张万里等, 2000),但并不表明次生林具有较强的稳定性(郝占庆等, 1994)。次生林的物种多样性和群落结构对不同程度的人为干扰如何响应,定量的研究还少见报道。
次生林的物种组成、群落结构、生物多样性状况与阔叶红松林有很大差异,而且低海拔、人为活动频繁区域,所遭受的干扰也更严重。研究次生林对不同的干扰水平的反应,并据此制定科学有效的森林经营和保育计划,对于次生林这种人为干扰形成的生态系统,尤其具有重要的意义(Zhang et al., 2000)。
本文对次生林和阔叶红松林的差异进行了比较,同时就干扰对次生林树种多样性和群落结构的影响进行了研究。
1 研究地点和研究方法长白山地处我国吉林省东部,山体东南坡与朝鲜接壤,最高峰海拔2 691 m,为我国东北最高山和3大河流发源地。长白山北坡为抚松、安图和和龙等县,植被垂直分异明显,其中海拔1 100 m以下为阔叶红松林带,海拔550 m以下已经成为次生的落叶阔叶混交林带(王战等, 1980;赵大昌, 1980)。
研究区年平均气温0.9~3.9 ℃,年均降水632.8~782.4 mm,为受季风影响的的温带大陆性山地气候,夏季温暖,冬季寒冷,降水集中在夏季。土壤为发育在火山灰沙砾土上的暗棕色森林土(徐振邦等, 1985)。
1.1 取样方法在长白山自然保护区、白河林业局,选取不同林型、不同立地条件、不同人为干扰水平的次生林林分设置样方。林场采伐作业区及公路、居民点附近等人为干扰较多的地点,海拔550~750 m,坡度0~10°。共计调查次生林样方49块,既有保护较好、多年未受人为干扰的林分,也有皆伐后天然恢复和新近受到采伐作业干扰的林分,样方面积20 m×30 m,于1995、1996年夏季进行调查。为了与次生林进行比较,2002年夏季还对长白山自然保护区内的阔叶红松原始林进行了调查,调查样方13块,样方面积20 m×30 m,分布于海拔700~1 000 m,坡度0~5°。调查中将每个样方划分为6个10 m×10 m的小样方。在每个小样方中对胸径≥3 cm的树木,记载种名,并测定胸径、树高。同时测定各样方的经纬度、海拔、坡度、坡向和坡位。
1.2 数据分析1) 物种多样性测度 物种丰富度:SR=物种数;Shannon指数: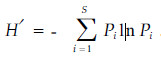
2) 逐步回归分析 干扰直接导致林分蓄积量和群落结构(如林分密度、胸径、树高)的显著变化,而这些指标与物种多样性关系密切(Condit et al., 1996; Aiba et al., 1999; Waide et al., 1999)。采用物种丰富度和Shannon指数分别与蓄积量、林分密度、平均胸径、最大胸径、平均树高、最大树高等指标进行逐步回归,以研究干扰对树种多样性的影响机制。
3) 方差分析 林分蓄积量的大小不仅直接受到干扰强度和干扰方式的影响,而且与干扰后的恢复时间的长短密切相关,是干扰方式、干扰强度和干扰历史的综合反映。为了定量评价干扰对物种多样性和群落结构的影响,本文以蓄积量作为衡量干扰水平的指标,采用方差分析研究不同的人为干扰水平对次生林树种多样性和群落结构的影响。
2 结果与分析 2.1 次生林和阔叶红松林的差异1) 树种组成的差异 次生林和阔叶红松林在树种组成上存在很大的差异:阔叶红松林主要由红松、紫椴(Tilia amurensis)、蒙古栎(Quercus mongolica)、臭冷杉(Abies nephrolepis)、水曲柳(Fraxinus mandshurica)等组成,平均RBA分别为32.0%、17.0%、10. 8%、10.5%、5.8%。次生林则主要由山杨(Populus davidiana)、蒙古栎、紫椴、黄榆(Ulmus macrocarpa)、白桦(Betula platyphylla)、核桃楸(Juglans mandshurica)等树种组成,平均RBA分别为20.4%、18.5%、16.8%、10.9%、8.3%、8.2%。与阔叶红松林相比,次生林的阳性树种占有很大比例,如山杨、黄榆、白桦、蒙古栎合计占次生林RBA组成的58%。尤其是山杨已经成为次生林中平均RBA最高的树种,即使在郁闭度较高(0.8 ~0.9),多年未受采伐的林分中也经常占有很高的比例(RBA可达30%~49%)。反映出次生林由于长期持续的人为干扰,有向旱生化方向逆行演替的明显趋势。另一个重要差别是,次生林中红松的平均RBA仅为0.19%。在所调查49个样方中,仅在接近保护区的6个样方中零星出现红松幼树,RBA仅0.2%~3.4%。其余样方均由于缺乏红松种源,未发现红松的天然更新幼苗。从物种组成上来看,长白山北坡的次生林林分质量较差,如果遭受进一步干扰,演替趋势不容乐观。
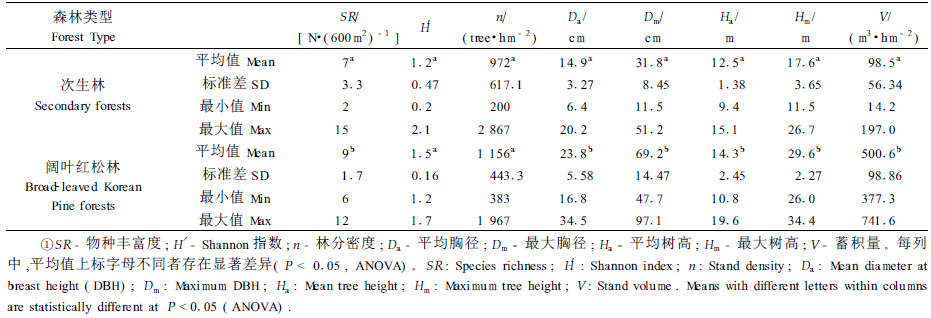
2) 树种多样性、群落结构和蓄积量的差异 从表 1可以看出,次生林的物种丰富度和Shannon指数都低于阔叶红松林,方差分析表明,次生林物种丰富度与阔叶红松林存在显著的差异。在蓄积量和群落结构指标中,除林分密度差异不显著外,其他指标均存在显著差异。阔叶红松林的平均胸径、最大胸径、最大树高和蓄积量平均值分别为23.8 cm、69.2 cm、29.6 m和500.6 m3·hm-2,次生林则分别仅为14.9 cm、31.8 cm、17.6 m和98.5 m3·hm-2,阔叶红松林分别为次生林的1.6、2.2、1.7和5.1倍。阔叶红松林蓄积量最高可达741.6 m3·hm-2,最低也有377.3 m3·hm-2。而次生林最高仅为197.0 m3·hm-2,最低可至14.2 m3·hm-2,可见次生林的林分质量十分低下。
|
|
1) 干扰对树种多样性影响的机制 从表 2可以看出,在次生林中,蓄积量能够显著解释物种丰富度和Shannon指数变化的51%和36% (P<0.01),而其他指标(胸径、树高)解释力很小。次生林的物种丰富度和Shannon指数都随蓄积量的减少而显著下降(表 2,图 1)。本研究的阔叶红松林样方位于保护区内,受到人为干扰很少,但由于自然干扰存在明显的林冠空隙动态(徐化成, 2001),蓄积量也存在较大的差异(表 1)。表 2表明,蓄积量可以显著解释阔叶红松林Shannon指数的变化(P<0.05),与次生林不同的是,Shannon指数随蓄积量的下降反而上升。
|
|

干扰直接导致各种群落结构和功能指标(如林分密度、胸径、树高和蓄积量)的变化,而树种多样性与森林群落的结构和功能密切相关(Aiba et al., 1999)。如林分密度的大小会直接影响物种多样性(Condit et al., 1996),林分生物量的不同也是影响物种多样性的一种重要机制(Waide et al., 1999)。本文的结果表明,不论是次生林还是阔叶红松林,干扰导致的树种多样性变化与蓄积量关系最为密切,而其他指标解释力很小(表 2)。如在次生林中,虽然林分密度与物种丰富度、Shannon指数之间存在显著的正相关(相关系数分别为0.69和0.46,P<0.01),但表 2表明林分密度并不能显著解释物种丰富度的变化,说明干扰导致的林分密度变化并不是导致树种多样性变化的主要原因。
蓄积量实际上反映了群落的生物量(Fang et al., 2001),在气候等环境梯度较小时,物种多样性与生物量一般呈驼峰形关系:生物量较小时,多样性随生物量的增加而提高,生物量较大时,多样性随生物量的增加而降低(Waide et al., 1999)。本文的结论与其一致,蓄积量较小的次生林属于前一种情况(图 1),阔叶红松林则属于后一种情况。阔叶红松林的树种多样性在适度的干扰下上升,与其他研究的结论一致(罗菊春等, 1997),也符合中度干扰假说(Connell, 1978)。一定的干扰如林冠空隙的产生会改变原始林内资源(如光照等)的分配,提供了更为丰富的生态位,使得更多的物种能够共存(Whitmore, 1989)。不过,次生林本身就是严重干扰的产物,中度干扰假说并不适用,其树种多样性随着干扰加剧而显著下降(图 1)。这种差异表明,次生林的树种多样性对人为干扰十分敏感,而阔叶红松林则有很强的抗干扰能力。

|
图 1 长白山北坡次生林样方的物种丰富度(a)、Shannon指数(b)与蓄积量的关系 Fig. 1 Relationship between species richness (a), Shannon index (b) of secondary forests and stand volume on northern slope of Mt. Changbai |
2) 干扰与群落结构的关系 由表 3、图 2可以看出,在次生林中,除平均胸径外所有群落结构指标均随蓄积量的下降而显著下降(P<0.01)。在阔叶红松林中,蓄积量仅与最大胸径在水平上显著正相关,其他指标均与蓄积量无显著的相关关系,说明阔叶红松林的自然干扰水平不足以造成多数群落结构指标的显著变化。这一结果表明,阔叶红松林的群落结构也有着很强的抗干扰能力,而次生林则对干扰十分敏感。
|
|


|
图 2 长白山北坡次生林样方的林分密度(a)、最大树高(b)与蓄积量的关系 Fig. 2 Relationship between stand density (a), maximum tree height (b) o f secondary forests and stand volume on northern slope of Mt. Changbai |
3) 不同干扰水平对次生林树种多样性和群落结构的影响 上述分析表明,蓄积量能够较好的反映次生林的树种多样性、群落结构在不同人为干扰水平下的变化,适宜作为衡量长白山北坡次生林干扰水平的指标。为了研究在何种人为干扰水平下次生林的多样性和群落结构发生显著变化,本文将蓄积量划分为4个水平表示不同程度的人为干扰(为了叙述方便,分别称为轻度、中度、强度和严重干扰),进行One-Way ANOVA方差分析,结果见表 4。可以看出次生林的平均胸径在不同的干扰水平上均无显著差异,但其余多样性和群落结构指标则有着显著的差异。
|
|

物种丰富度在轻度和中度干扰下无显著变化,但在强度和严重干扰下出现持续、显著的下降。Shannon指数在严重干扰下与轻度干扰之间存在显著的差异。可以看出,强度的干扰(蓄积量<100 m3·hm-2)即对次生林树种多样性造成了显著的影响。
在群落结构指标中,林分密度在强度、严重干扰下与轻度干扰之间存在显著差异。最大胸径在严重干扰下显著下降。平均树高对干扰最为敏感,在中度干扰水平就显著下降。最大树高则在强度及严重干扰下出现与轻度干扰的显著差异。这一结果表明,中度的干扰(蓄积量<150 m3·hm-2)对群落结构即开始产生显著影响,而强度的干扰(蓄积量<100 m3 ·hm-2)则足以造成多数群落结构指标的显著下降。
3 结论与讨论对长白山北坡次生林和阔叶红松林的比较表明,二者在物种组成、多样性和群落结构上都存在很大的差异。次生林阳性树种所占比例过高,红松所占比例相当低,由于缺乏种源,红松天然更新不良。同时,次生林的树种多样性、群落结构指标和蓄积量均显著低于阔叶红松林,其中蓄积量仅为阔叶红松林的20%(表 1),林分质量和生产力十分低下。
对树种多样性的分析表明,干扰导致的蓄积量变化是树种多样性变化的主要原因,而林分密度、胸径、树高等指标的变化对多样性影响很小。其中,阔叶红松林的多样性在适度自然干扰下有所上升,而次生林树种多样性则随着人为干扰的加剧而显著下降(表 2,图 1)。对群落结构的分析表明,阔叶红松林的群落结构指标在自然干扰下未发生显著的变化,而次生林则随着人为干扰的加剧而显著下降(表 3,图 2)。这些结果反映出阔叶红松林有很强的抗干扰能力,而次生林不论是树种多样性还是群落结构对干扰都十分敏感。
虽然国内外就干扰对森林群落结构和生物多样性的影响已经有了较多的研究,但对干扰水平的定量评价一直比较困难。然而,如果不对干扰水平进行定量的评价,在何种干扰下森林群落的结构、组成和多样性会发生显著的变化等问题就无法确定。本文尝试采用蓄积量作为衡量干扰水平的指标。蓄积量是干扰强度、干扰方式、干扰历史综合作用的结果,既能够反映干扰的强度,也能反映干扰后林分的恢复情况;而且其测定简便易行,在生产实践中应用十分广泛。文中的分析也表明,蓄积量能够较好的反映长白山北坡次生林在不同人为干扰下树种多样性和群落结构的变化(表 2、3,图 1、2),可以作为衡量次生林干扰水平的指标。对不同干扰水平下次生林树种多样性和群落结构指标的分析表明(表 4),中度的干扰(蓄积量<150 m3·hm-2)就会导致群落结构的显著变化,而强度的干扰(蓄积量<100 m3 ·hm-2)已足以导致次生林的多样性和群落结构指标的严重下降。
综上所述,长白山北坡次生林已经出现明显的逆行演替的趋势, 其多样性、林分质量和生产力十分低下,对人为干扰十分敏感,而且多数地方缺乏红松种源。He等(2002)对长白山保护区及其附近森林演替的模拟研究表明,如果仅依赖红松种源的天然扩散,即使在没有任何人为干扰的情况下,从次生林向阔叶红松林的恢复至少需要500~600年。因此只有加强对次生林的保护,同时采取栽针保阔的措施(周晓峰, 1982),才能遏制目前次生林逆行演替的趋势,促进其向阔叶红松林方向恢复。
郝占庆, 陶大立, 赵士洞. 1994. 长白山北坡阔叶红松林及其次生白桦林高等植物物种多样性比较. 应用生态学报, 5(1): 16-23. DOI:10.3321/j.issn:1001-9332.1994.01.013 |
郝占庆. 2000.长白山北坡植物群落多样性及其梯度格局分析.博士论文.沈阳: 中国科学院沈阳应用生态研究所
|
雷相东, 张会儒, 李冬兰, 等. 2003. 东北过伐林区四种森林类型的物种多样性比较研究. 生态学杂志, 22(5): 47-55. DOI:10.3321/j.issn:1000-4890.2003.05.011 |
罗菊春, 王庆锁, 牟长城, 等. 1997. 干扰对天然红松林植物多样性的影响. 林业科学, 33(6): 498-503. DOI:10.3321/j.issn:1001-7488.1997.06.003 |
王战, 徐振邦, 李昕, 等. 1980. 长白山北坡主要森林类型及其群落结构特点. 森林生态系统研究, 1: 1-8. |
吴方正. 2003. 东北阔叶红松林分布区生态气候适宜性及全球气候变化影响评价. 应用生态学报, 14(5): 771-775. DOI:10.3321/j.issn:1001-9332.2003.05.028 |
徐化成. 2001. 中国红松天然林. 北京: 中国林业出版社.
|
徐振邦, 李昕, 戴洪才, 等. 1985. 长白山阔叶红松林生物生产量的研究. 森林生态系统研究, 5: 33-47. |
徐振邦. 1985. 合理经营中国温带阔叶红松林提高林地生产力的研究. 森林生态系统研究, 5: 57-63. |
阳含熙, 谢海生. 1994. 长白山红松混交林干扰历史的重构研究. 植物生态学报, 18(3): 201-208. DOI:10.3321/j.issn:1005-264X.1994.03.007 |
臧润国. 1995.红松阔叶林林冠空隙动态的研究.博士论文.北京: 北京林业大学 http://cdmd.cnki.com.cn/Article/CDMD-10022-2006106785.htm
|
张万里, 李雷鸿. 2000. 黑龙江省东部林区森林植物生物多样性与干扰的研究. 东北林业大学学报, 28(5): 77-82. DOI:10.3969/j.issn.1000-5382.2000.05.018 |
赵大昌. 1980. 长白山的植被垂直分布带. 森林生态系统研究, 1: 65-70. |
周晓峰. 1982. 红松阔叶林的恢复途径——栽针保阔. 东北林学院学报, 10(增刊): 18-28. |
Aiba S I, Kitayama K. 1999. Structure, composition and species diversity in an altitude_substrate matrix of rain forest tree communities on Mount Kinabalu, Borneo. Plant Ecology, 140: 139-157. DOI:10.1023/A:1009710618040 |
Condit R, Hubbell S P, Lafrankie J V, et al. 1996. Species_area and species_individual relationships for tropical trees: a comparison of three 50_ha plots. Journal of Ecology, 84(4): 549-562. DOI:10.2307/2261477 |
Connell J H. 1978. Diversity in tropical rain forests and coral reefs. Science, 199: 1302-1310. DOI:10.1126/science.199.4335.1302 |
Fang J Y, Chen A P, Peng C H, et al. 2001. Changes in forest biomass carbon storage in China between 1949 and 1998. Science, 292: 2320-2322. DOI:10.1126/science.1058629 |
He H S, Hao Z, Larsen D R, et al. 2002. A simulation study of landscape scale forest succession in northeastern China. Ecological Modelling, 156: 153-166. DOI:10.1016/S0304-3800(02)00104-7 |
Waide R B, Willig M R, Steiner C F, et al. 1999. The relationship between productivity and species richness. Annual Review of Ecology and Systematics, 30: 257-300. DOI:10.1146/annurev.ecolsys.30.1.257 |
Whitmore T C. 1989. Canopy gaps and two major groups of forest tree. Ecology, 70(3): 536-538. DOI:10.2307/1940195 |
Zhang P, Shao G, Zhao G, et al. 2000. China's Forest Policy for the 21st Century. Science, 288: 2135-2136. DOI:10.1126/science.288.5474.2135 |