 2006, Vol. 42
2006, Vol. 42文章信息
- 张萍, 周志春, 金国庆, 范辉华, 胡红宝.
- Zhang Ping, Zhou Zhichun, Jin Guoqing, Fan Huihua, Hu Hongbao.
- 木荷种源遗传多样性和种源区初步划分
- Genetic Diversity Analysis and Provenance Zone Allocation of Schima superba in China Using RAPD Markers
- 林业科学, 2006, 42(2): 38-42.
- Scientia Silvae Sinicae, 2006, 42(2): 38-42.
-
文章历史
- 收稿日期:2004-03-19
-
作者相关文章



2. 福建省建瓯市林业科技推广中心 建瓯 353100;
3. 浙江省淳安县林业局 淳安 311700
2. The Extending Center for Forestry Science and Technology of Jian'ou city, Fujian Province Jian'ou 353100;
3. Forestry Bureau of Chun'an County, Zhejiang Province Chun'an 311700
利用DNA标记可以直接在分子水平上研究物种的遗传多态性。在多种分子标记中,RAPD具有多态性高、所需DNA量少、取样方便、能实现全基因组无偏取样和无组织器官特异性等优点,已应用于遗传多样性检测、QTLs定位、品系鉴定、遗传图谱构建和生物系统学等研究,在群体遗传结构和遗传多样性研究方面应用尤为广泛。如Wachira等(1995)和Kaundun等(2000)利用RAPD标记系统研究了茶树(Camellia sinensis)的遗传多样性。苏晓华等(1996)基于RAPD标记揭示了大青杨(Populus ussuriensis )及其近缘种的遗传变异和亲缘关系。
木荷(Schima superba)属山茶科(Theaceae)木荷属常绿乔木,为我国东部湿润亚热带常绿阔叶林的重要成分之一,自然分布于北纬31°以南,东经105°以东的广大地区,是我国南方山区重要的高效生物防火和优良用材树种(倪健, 1996)。由于木荷自然分布区广泛,种内变异丰富,针对以高效生物防火和优质用材为目标的木荷育种对于我国南方生态公益林快速构建和商品用材林建设意义重大。但有关木荷遗传多样性的研究较少,仅见王峥峰等(2000)利用AFLP标记对广东鼎湖山3个不同演替系列木荷群落遗传多样性的研究,发现95.99%的遗传变异表现在群落内,4.01%的遗传分化表现在群落间。作者基于7省区36个木荷种源的苗期测定结果,发现木荷苗期主要经济性状存在显著的种源差异,且多呈典型的纬向变异模式,速生种源主要来自25°N左右的自然分布区(张萍等, 2004)。本文则进一步利用RAPD分子标记研究木荷不同地理种源的遗传多样性和种源间的遗传分化,旨在为木荷育种策略的科学制定和遗传资源的有效保护提供理论依据。
1 材料与方法 1.1 试验材料试验叶样取自浙江省桐庐县的木荷种源苗期试验2年生留床苗。取样种源为来自7省区的15个代表性种源,包括安徽太平、浙江临海、浙江龙泉、湖南桑植、湖南浏阳、湖南茶陵、湖南嘉禾、江西铜鼓、江西永丰、福建尤溪、福建武平、广东韶关、广东广宁、广西桂林和广西梧州。木荷种源试验的采种林分为当地起源的天然纯林或混交林,年龄15年以上,林相整齐,分布较均匀,结实较多。选择20株以上的优良母树采种,母树间距50 m以上。参试种源的基本情况和苗期遗传测定等详见张萍等(2004)一文。2003年5月下旬,每个种源随机抽取12个单株,分别采集其顶端嫩叶用于DNA提取。
1.2 总DNA提取每个单株取新鲜嫩叶4 g,采用CTAB法提取DNA(Doyle et al., 1987)。对提取的DNA利用日本岛津UV-2401PC紫外分光光度计测定260 nm/280 nm处的光吸收值来检测其纯度和浓度,再用1.2%的琼脂糖凝胶电泳进行检测。
1.3 引物筛选选择8个代表性的DNA样品进行引物筛选,从150个随机引物筛选15个能产生多态、清晰且可重复条带的引物,然后利用这些引物检测所有样品。各引物名称、序列及检测的位点总数和多态性位点数见表 1。
|
|

RAPD扩增反应在Perkin_Elmer 9600基因扩增仪上进行。RAPD扩增反应条件为:94 ℃预变性3 min;然后进入45个循环,每个循环94 ℃变性30 s,37 ℃退火30 s,72 ℃延伸2 min;循环完成后为保证DNA延伸充分再72 ℃延伸7 min,最后于4 ℃下保存。PCR反应采用20 μL体系:10倍反应缓冲液2 μL(100 mmol·L-1 Tris_HCl,pH9.0,500 mmol·L-1 KCl,Mg2+ 2 μL 25 mmol·L-1),0.01%明胶,5.0 g·L-1 BSA、Taq酶1/3 μL(1 Unit)、dNTP 2 μL(终浓度0.1 mmol·L-1)、引物1 μL(稀释为5 μmol·L-1)、DNA模板5 ng。扩增产物经1.5%琼脂糖凝胶电泳分离,再经溴化乙锭染色,最后利用FR-200紫外可见光分析成像系统拍照记录结果。图 1给出了引物S141的RAPD扩增产物在部分木荷种源植株样品中的分离情况。

|
图 1 引物S141的RAPD扩增产物在部分木荷样品中的分离 Fig. 1 Segregation of RAPD bands amplified by S141 in some sample of S. superba |
RAPD带按1/0标记,利用POPGENE(Tools for Population Genetic Analysis)软件(Version 1.3.1)进行数据分析,计算多态位点百分率、Shannon表型多样性指数( I)、总的种源基因多样度(HT)、种源内基因多样度(HS)、种源间基因多样度(DST)、基因分化系数(GST)、基因流(Nm)等以衡量木荷种源的遗传多样性和遗传分化。最后利用RAPDistance软件计算出所有个体遗传相似系数,联合WINAMOVA1.55(Analysis of Molecular Variance)分析软件(Excoffier et al., 1992)进行分子方差分析,计算群体间和群体内的方差分量并作显著性检验,以此度量群体间和群体内遗传变异水平。最后基于种源间Nei's无偏遗传距离(Unbiased genetic distance)的估算(Nei, 1978),对15个种源进行UPGMA聚类分析。
2 结果与分析 2.1 RAPD多态性用15个10 bp的随机引物对来自15个木荷种源的180个个体的基因组DNA进行RAPD分析,共扩增出86条清晰且可重复的带,大小为250~2 100 bp。扩增结果发现,各引物检测到的RAPD位点数介于4~8,多态位点为2~6个。在检测的86个位点中,70个位点呈多态性,多态位点百分率为81.40%。表 2给出的结果表明,木荷种内多态位点百分率差异很大,福建尤溪种源的多态位点百分率最高,达78.61%,而安徽太平种源最低,为54.35%,前者是后者的1.45倍。
|
|
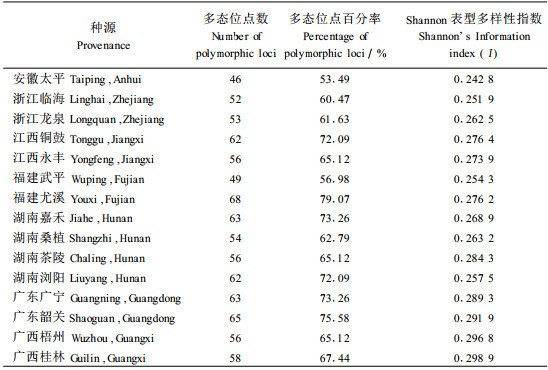
这里利用Shannon表型多样性指数来说明木荷遗传多样性的种源差异(表 2)。结果表明木荷这一广域性树种不同种源间遗传多样性相差很大,种源的Shannon表型多样性指数变化在0.242 8~0.298 9之间。如分布区北部的安徽太平种源Shannon表型多样性指数为0.242 8,而分布区南部的广西桂林种源为0.298 9,是安徽太平种源的1.23倍。与产地经纬度的简单相关分析发现,木荷种源Shannon表型多样性指数与产地纬度呈显著的负相关,相关系数-0.580 5(p=0.040 2),而与产地经度则有一定程度的正相关,相关系数为0.465 7(p=0.010 9)。这说明来自木荷自然分布区南部的种源具有较高的遗传多样性,而北部种源的遗传多样性则较低,中西部种源的遗传多样性有比东部种源低的趋势。
2.3 种源间的遗传分化表 3结果显示,木荷种内平均基因多样度为0.363 6,种源内平均基因多样度为0.265 1,种源间的基因多样度为0.098 5。基因分化系数达到0.271 4,基因流较小,仅为0.671 1。虽然木荷为我国亚热带地区广域性分布的地带性树种,但因其为虫媒异花授粉,种源间的基因流与松杉等风媒授粉树种相比较小,种源间的遗传分化较大,27.14%的遗传变异存在于地理种源间,而种源内的变异占到了总变异的72.86%。
|
|

AMOVA分析结果(表 4)亦表明,木荷种源间和种源内都存在显著的遗传变异(p<0.01),其中种源间变异占28.78%,种源内变异占71.22%。这一结果与利用POPGENE软件分析获得的结论基本一致。
|
|

估算15个木荷种源间Nei's无偏遗传距离(Nei, 1978),并据此使用UPMGA聚类法得出树状图(图 2)。估算结果发现,木荷种源间的遗传距离在0.005 7~0.335 3,广东韶关和湖南嘉禾种源间的遗传距离最小(0.005 7),两种源在地理距离上也很近;广东广宁和安徽太平种源间的遗传距离最大(0.335 3),两者在地理距离上也很远。图 2聚类树状图显示,在DNA水平上可以较好地将木荷划分为3个种源区。第1种源区为南部种源区,包括广东广宁、韶关,广西桂林、梧州,湖南嘉禾、茶陵和福建武平种源,这些种源大致分布在25°N左右的地区,也即南岭山脉等地区。其中广东广宁、韶关,广西桂林、梧州,湖南嘉禾这5个在地理上相邻的种源,地理亲缘关系密切,其间遗传距离在0.005 7~0. 109 2,平均为0.081 8,而福建武平和湖南茶陵则与上述5个种源的遗传距离较远,因此可将南部种源区进一步分为东部和西部亚区。第2类为中部种源,包括浙江龙泉,江西铜鼓、永丰,福建尤溪,湖南浏阳、桑植等,这些种源分布于南岭以北、浙江南部以南等广大地区,其间遗传距离在0.057 5~0.187 7,平均为0.140 9。较之于南部种源,木荷中部分布区相邻种源间的遗传分化较大,其间的平均遗传距离为南部种源间遗传距离的1.72倍。从图 2可看出,地处木荷自然分布区西部的湖南桑植种源与中部的其他种源亲缘关系较远,平均遗传距离0.131 3,因此,同样可将木荷中部种源再分为东部和西部2个亚区。第3类即为木荷的北缘种源,包括安徽南部和浙江北部种源。

|
图 2 15个木荷种源的UPGMA聚类图 Fig. 2 Dendrogram resulting from UPGMA analysis of 15 provenances of S. superba from China based on pairwise values for Nei's (1978) unbiased genetic distance |
对于缺少遗传背景研究的树种,通过系统研究其种内遗传多样性和遗传结构,可为制定科学的遗传改良和保育策略提供理论基础。作为我国亚热带地区高效生物防火和优质用材树种,木荷在生态公益林和商品用材林建设中发挥了重要作用。作者基于木荷苗期性状种源变异和地理模式的研究,已证实其种源间差异的显著性(张萍等, 2004),这对于以高效生物防火和优质用材为不同培育目标的优良种源选择奠定了重要理论基础。然而有关木荷种群遗传结构和遗传多样性的研究较少。王峥峰等(2000)仅从分子生态学角度,利用AFLP标记揭示了广东鼎湖山自然保护区3个不同演替系列木荷群落遗传多样性的变化,研究发现木荷种群基因多样度为0.331,不同演替阶段木荷种群的遗传分化很小,种群间遗传变异仅占总变异的4.01%,而种群内的变异则占了总变异的95.99%。
本文利用RAPD标记研究了木荷15个代表性地理种源的遗传多样性。研究发现木荷遗传多样性丰富,其多态位点百分率高达81.40%,明显高于韦塔桉( Eucalyptus wetarensis)的77.0%(潘天玲等, 2003)和马褂木的41.04%(李建民等, 2002)。木荷种内平均基因多样度为0.363 6,不同种源的Shannon表型多样性指数差异巨大,以分布区北部的安徽太平种源最小(0.242 8),分布区南部的广西桂林种源最大(0.298 9)。木荷种源Shannon表型多样性指数与产地纬度则呈显著的负相关,与产地经度有一定程度的正相关,来自分布区北部的种源遗传多样性较低,而南部种源尤其是来自南岭山脉地区(也即25°N左右区域)的种源遗传多样性较高。南岭山脉地区水热资源优越,是我国生物多样性和遗传多样性最丰富的区域之一。尤勇等(1998)在对杉木(Cunninghamia lanceolata)种源遗传多样性研究的基础上,认为南岭山脉西部是杉木中心产区之一。根据木荷种源苗期试验(张萍等, 2004)和Shannon表型多样性指数估算结果,可以推测25°N左右的南岭山脉附近可能是木荷的中心产区。研究发现,木荷约有30%的遗传变异来源于其地理种源间,70%左右的变异存在于地理种源内。
种源区的科学划分是制定林木种子合理调拨原则的基础。通常利用种源多点区域试验结果划分种源区,耗用时间、经费和人力巨大(徐化成, 1990)。在未进行种源试验,或种源试验结果还未获得之前,可利用分子标记直接在DNA水平进行种源聚类,以此作为辅助对种源区进行初步划分,这样所花时间和费用将大大减少。本文在木荷种源遗传多样性和种源遗传分化研究的基础上,可较好地将木荷分布区划分为3个种源区,即北缘种源区(安徽南部和浙江北部)、中部种源区(南岭以北、浙江南部以南)和南部种源区(南岭以南),而南部种源区和中部种源区又都可再分为东部和西部2个种源亚区。
李建民, 周志春, 吴开云, 等. 2002. RAPD标记研究马褂木地理种源的遗传分化. 林业科学, 38(4): 61-66. DOI:10.3321/j.issn:1001-7488.2002.04.010 |
倪建. 1996. 中国木荷与木荷林的地理分布与气候的关系. 植物资源与环境, 5(3): 28-34. |
潘天玲, 刘友全. 2003. RAPD标记在韦塔桉种源遗传结构上的应用. 生命科学研究, 7(1): 89-94. DOI:10.3969/j.issn.1007-7847.2003.01.017 |
苏晓华, 张绮纹, 张望东, 等. 1996. 大青杨及其近缘种的遗传变异和系统关系研究. 林业科学, 32(2): 118-124. |
王峥峰, 王伯荪, 李鸣光, 等. 2000. 南亚热带森林优势种源荷木和锥栗在演替系列群落中的分子生态研究. 植物学报, 42(10): 1082-1088. DOI:10.3321/j.issn:1672-9072.2000.10.016 |
徐化成. 1990. 林木种子区划. 北京: 中国林业出版社.
|
尤勇, 洪菊生. 1998. RAPD标记在杉木种源遗传变异上的应用. 林业科学, 34(4): 32-38. DOI:10.3321/j.issn:1001-7488.1998.04.005 |
张萍, 金国庆, 周志春, 等. 2004. 木荷苗木性状的种源变异和地理模式. 林业科学研究, 17(2): 192-198. DOI:10.3321/j.issn:1001-1498.2004.02.009 |
Doyle J J, Doyle J L. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf material. Phytochem Bull, 19: 11-15. |
Excoffier L, Smouse P E, Quattro J M. 1992. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochoudrial DNA restriction data. Genetics, (131): 479-491. |
Kaundun S S, Zhyvoloup A, Park Y G. 2000. Evaluation of genetic diversity among elite tea (Camellia sinensis var. sinensis)accessions using RAPD markers. Euphytica, 115: 7-16. |
Nei M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, (89): 583-590. |
Wachira F N, Waugh R, Hackett C A, et al. 1995. Detection of genetic diversity in tea (Camellia sinensis) using RAPD markers. Genome, 38(2): 201-210. DOI:10.1139/g95-025 |