 2006, Vol. 42
2006, Vol. 42文章信息
- 刘兴良, 刘世荣, 宿以明, 蔡小虎, 马钦彦.
- Liu Xingliang, Liu Shirong, Su Yiming, Cai Xiaohu, Ma Qinyan.
- 巴郎山川滇高山栎灌丛地上生物量及其对海拔梯度的响应
- Aboveground Biomass of Quercus aquifolioides Shrub Community and Its Responses to Altitudinal Gradients in Balangshan Mountain, Shichuan Province
- 林业科学, 2006, 42(2): 1-7.
- Scientia Silvae Sinicae, 2006, 42(2): 1-7.
-
文章历史
- 收稿日期:2004-05-31
-
作者相关文章



2. 四川省林业科学研究院 成都 610081;
3. 中国林业科学研究院森林生态环境与保护研究所 北京 100091
2. Sichuan Academy of Forestry Chengdu 610081;
3. Institute of Forest Ecology, Environment and Protection, CAF Beijing 100091
川滇高山栎(Quercus aquifolioides)林属亚高山硬叶栎林(sub_alpine sclerophyllous oak forest)(杨钦周,1990),分布于川滇两省交界处金沙江河谷W形弯曲处,其邻近地区种群强烈分化,形成向东西扩展的趋势,其垂直分布范围极广,生态适应性很强,在横断山地区广泛分布,随海拔由南向北逐渐升高(徐润清等,1992; 周立江等,1992; 周浙昆,1993),出现了从森林到灌丛连续分布的罕见现象(中国植被编辑委员会,1980; 姜汉侨,1980; 金振洲,1979; 1987;金振洲等,1981; 李文华,1989)。川滇高山栎灌丛经济价值不高,但根系发达,萌生能力强,适应范围广,在西南山地有良好的水土保持和水源涵养作用(杨玉坡等,1992)。森林生物量和生产力是整个生态系统运行的营养物质来源和能量基础,也是判断大气中CO2源和汇的重要标志(Houghton et al., 1983; 1985;1987;Grubler, 1993; Iverson et al., 1993; 1994;冯宗炜等,1999)。青藏高原森林生物量和生产力的相关研究资料极为匮乏(冯宗炜等,1999),研究种群的地理分化以及种群内个体形态分化成了重点发展方向(方精云等,1993; 钟章成等,2001)。因此,系统研究川滇高山栎灌丛群落结构与功能、生物量与生物生产力及其对环境的响应,可以对川滇高山栎灌丛林的评价、合理经营及充分发挥其综合效益提供理论依据,也可以为川滇高山栎灌丛的起源及其形成机制、能量流动与养分循环等积累基础资料。
1 自然概况卧龙自然保护区位于青藏高原东南缘的邛崃山东坡(102°52′—103°24′ E,30°45′— 31°25′ N),以高山峡谷为主要地貌特征。该区冬寒夏凉、降水丰富、干湿季节明显,在海拔2 700 m地带,年平均温度8.4 ℃,1月平均气温-1.7 ℃,7月平均气温17.0 ℃,≥0 ℃活动积温4 000 ℃; 年平均降水量861.8 mm,5—9月降水量占全年降水量的68.1%,年平均相对湿度80%左右。川滇高山栎灌丛集中分布在该区海拔2 700 ~3 300 m地带,下接河谷,上接高山草甸。林下土壤为山地棕壤,较干燥,厚度50 cm左右。灌丛郁闭度0.8,平均高度1.1~3.5 cm,以川滇高山栎为优势建群种,伴生种类主要有木帚栒子(Cotoneaster dielsianus)、平枝栒子(Cotoneaster horizontalis)、鞘柄菝葜(Smilax stans)、红花蔷薇( Rosa moyesii)等; 草本植物32种,盖度0.4,平均高度0.25 m,主要种类有糙野青茅(Deyeuxia scabrescens)、双花堇菜(Viola biflora)、珠芽蓼(Polygonum viviparum)、乳白香青( Anaphalis lactea)、钉柱委陵菜(Potentilla saundersiana)等; 苔藓盖度0.4,厚度2 cm; 枯枝落叶层厚度3~6 cm。
2 研究方法 2.1 野外调查在卧龙自然保护区巴郎山,沿川滇高山栎灌丛分布的5个不同海拔高度和不同生境设置5 m×5 m样方18个,每个海拔带依次相差100 m。在样方内实行每株检尺,测定离地10 cm处直径(D)及林木高度(H)。在每个海拔地带中选1丛,采用“分层切割法”(Monsi, 1968; Fujimori et al., 1976; 冯宗炜等,1982)按不同径级和高度实测川滇高山栎单株的干、皮、枝、叶器官生物量并取样称鲜质量,共计6丛24株。采用样方收获法(木村允,1976),在每个样方的四角和中心各设置小样方测定灌木、草本、苔藓和枯枝落叶生物量并取样称鲜质量,伴生灌木采用2 m×2 m样方测定枝干、叶生物量,草本采用1 m×1 m样方测定地上部生物量,苔藓和枯枝落叶采用0.5 m×0.5 m样方进行生物量测定,同时取样称鲜质量。
2.2 室内处理及分析将野外样品在试验室烘箱中90 ℃烘2 h后,85 ℃烘至恒重,用电子天平称质量,求样品干鲜质量比,将各器官鲜质量换算成干质量,根据样方资料换算单位面积干质量生物量。
2.3 分析与建模川滇高山栎生物量统计模型的建立与筛选在SPSS Base 11.5软件上进行; 用实测的地上部生物量及各器官生物量,先分析自变量参数与各器官生物量的相关性(刘庆,1999),然后依据相对生长(allometric relative growth)法则(Huxley,1931),应用CAR (constant allometric ratio)模型(Kittedge, 1944)和VAR(variable allometric ratio)模型(Ruark et al., 1987),筛选生物量统计模型,计算川滇高山栎地上部分生物量。
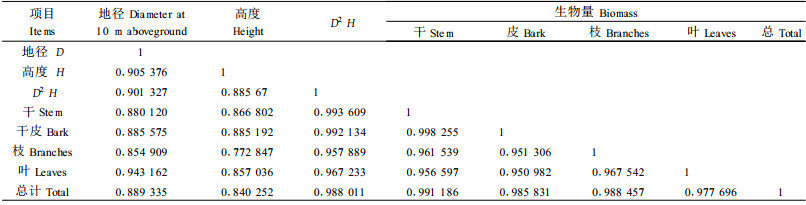
3 结果与分析 3.1 生物量模型的建立与选择依据相对生长法,对川滇高山栎灌丛各径级个体地径、植株高度与各器官生物量进行相关显著性分析。结果表明(表 1),川滇高山栎灌丛个体地径、植株高度与各器官生物量间具有显著的相关,且川滇高山栎个体地径、植株高度、以及干、皮、枝、叶、总生物量等相互间的相关均达到极显著水平。以生物量(W)为因变量,地径(D)、树高(H)、D2 H为自变量, 采用Y=aXb, Y=aeb, Y=aX+b, Y= 1/(u+abX), Y=a+blnX等数学模型进行回归拟合,求出相应参数a, b的值,经筛选建立了估测川滇高山栎单株器官生物量的统计模型(表 2)。从筛选建立的川滇高山栎单株器官生物量的统计模型来看,用地径(D)、树高(H)估测单株林木器官生物量的适合模型均为指数模型和幂函数模型,以指数模型为最佳,相关系数为0.982~0.996;而用D2H估测单株林木器官生物量的适合模型均为直线和指数模型,以直线统计模型为最佳,指数模型也能达到估测效果,相关系数为0.882~0.996。从表 2还可以看出,所筛选统计模型的相关系数都较高,并达到极显著水平,而且剩余回归标准差较小,说明估测值与实测值的拟合度高,估测误差小; 同时,该组统计模型消除了树种、林型和立地条件不同的影响,可见所筛选的统计模型估测林木生物量的效果较好。在实际应用中,选用地径(D)或树高( H)可估测川滇高山栎单株生物量,根据样地的每木资料即可计算川滇高山栎灌木的单位面积生物量。
|
|
|
|
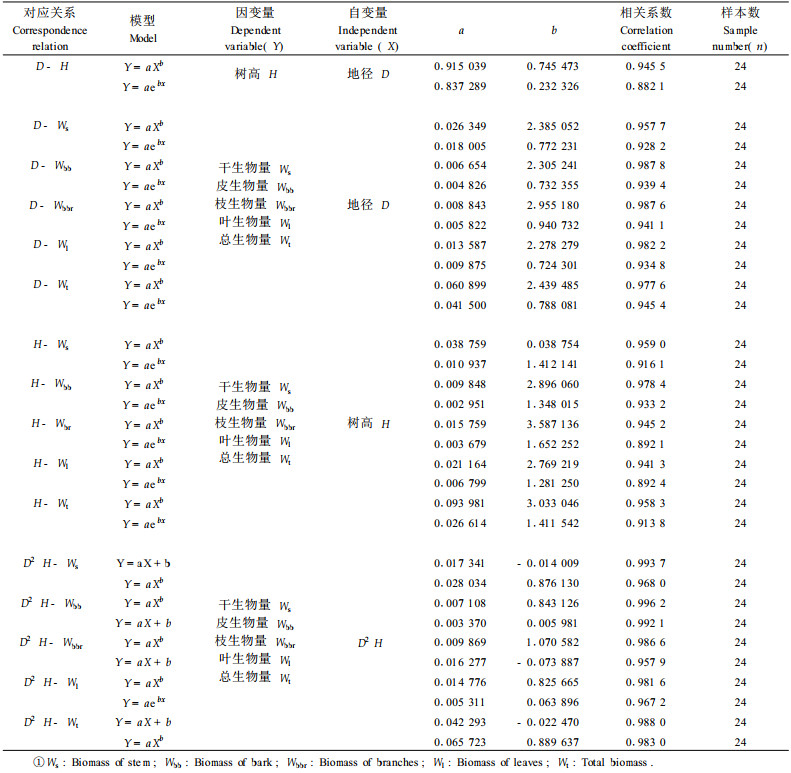
研究结果表明(表 3),川滇高山栎灌丛群落地上部生物量占绝对优势,平均为25.22 t·hm-2,各层次生物量的大小为:川滇高山栎>枯枝落叶层>伴生灌木>苔藓层>草本层。川滇高山栎生物量为18.21 t·hm-2,占总生物量的72.20%;枯枝落叶层生物量为5.98 t·hm-2,占总生物量的23.71%;伴生灌木的生物量为0.46 t·hm-2 ,占总生物量的1.80%;苔藓层生物量为0.42 t·hm-2,占总生物量的1.66%;草本层生物量最低为0.16 t·hm-2,占总生物量的0.63%。
|
|
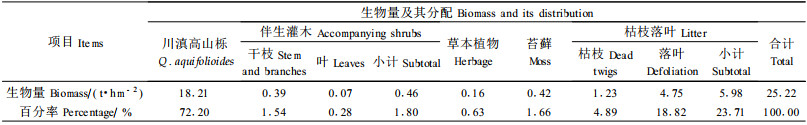
川滇高山栎种群地上部生物量为18.21 t·hm-2,各器官生物量大小为:树干>枝>叶>皮。其中干的生物量为7.88 t·hm-2,占总生物量的43.28%;枝生物量为4.90 t·hm-2,占总生物量的26.88%;叶生物量为3.61 t·hm-2,占种群总生物量的19.82%;皮生物量最低的为1.83 t·hm-2,仅占种群总生物量的10.02%(表 4)。
|
|
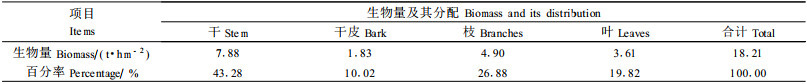
川滇高山栎灌丛内虽然伴生灌木种类较多,但密度小,且多数种类矮小,故地上部生物量不高,仅为0.46 t·hm-2,其中干枝和叶的生物量分别为0.39和0.07 t·hm-2,各占川滇高山栎灌丛地上部总生物量的1.54%和0.28%,各占伴生灌木地上部总生物量的84.78%和15.22%(表 3)。
3.2.4 枯枝落叶层生物量及其分配川滇高山栎为常绿树种。研究表明:树枝上除保留当年生叶之外,上年生叶约保留25%,故每年的落叶量较大,其总生物量为5. 98 t·hm-2,其中落叶大于枯枝,前者生物量为4.75 t·hm-2,占枯枝落叶总量的79.43%,占川滇高山栎灌丛地上生物量的18.82%;后者为1.23 t·hm-2,占枯枝落叶总量的20.57%,占川滇高山栎灌丛地上生物量的4.89%(表 3)。
3.3 川滇高山栎种群生物量对不同海拔梯度的响应从5个海拔高度川滇高山栎种群生物量的测定结果看(图 1),川滇高山栎地上部生物量随着海拔升高而降低。在海拔2 920 m以下,川滇高山栎灌丛群落生物量差异不大。海拔3 020 m处生物量下降非常明显,2个海拔高度总生物量差异达3倍以上。海拔2 720 m处与海拔3 120 m草甸接壤处川滇高山栎灌丛生物量差异达5倍以上。这是由于海拔高度不同,光、热、水、气组合产生差异,导致气温随着海拔的升高而降低,本区川滇高山栎分布幅度达1 000 m以上(即分布海拔2 200~3 600 m),由于本区域海拔每升高100 m,气温降低0.56 ℃(杨玉坡等,1992),因此,温差可达5.6 ℃以上,尤其是川滇高山栎灌丛与高山草甸的接壤地带,气候更加恶劣,植物生长期缩短,有效积温降低,影响了植物有机物的形成(江洪等,1985)。虽然灌丛密度和直径差异不大,但植株平均高度差异较大,导致了川滇高山栎灌丛种群生物量的差异。在海拔2 720~3 020 m地带,川滇高山栎地上部分各器官生物量的比例和排序具有一定的规律性(表 5),而且同高度川滇高山栎灌丛地上部分器官生物量大小为:干>枝>叶>皮,在海拔2 720 m处其比例为4.38:2.93:1.96:1.00;海拔2 820 m处比例为4.34:2.77:1.97:1.00;海拔2 920 m处比例为4.36:2.86:1.97:1.00;海拔3 020 m处比例为4.27:2.47:2.00:1.00;而海拔在3 120 m处比例为4.12:1.91:2.03:1.00。可以看出,川滇高山栎地上部分器官生物量的比例总趋势为:在海拔2 720~2 920 m,干:枝:叶:皮的比例约为4:3:2:1;在海拔3 120 m及3 020 m处,干:枝:叶:皮的比例约为4:2:2:1。

|
图 1 不同海拔高度川滇高山栎种群生物量的变化 Fig. 1 Variety of Q. aquifolioides bia mass at different altitude |
|
|

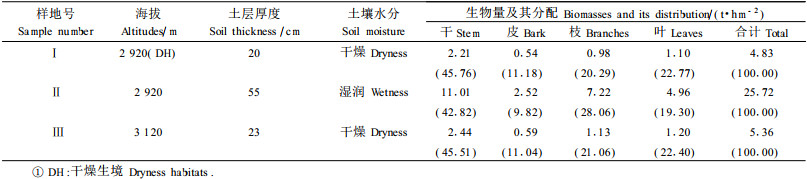
选择海拔2 920 m的2种不同生境与海拔3 120 m灌丛草甸带的川滇高山栎灌丛进行比较分析,结果为(表 6):同一海拔地带的2个样地比较,Ⅰ样地土层相对较薄,土壤干燥,Ⅱ样地土层相对较厚,湿度较大,其直径和高度均有很大差异。Ⅱ样地川滇高山栎灌丛生物量为25.72 t·hm-2,是样地Ⅰ的5.33倍。海拔2 920 m地带立地条件差的Ⅰ样地川滇高山栎灌丛总生物量与海拔3 120 m灌丛草甸带的Ⅲ样地川滇高山栎灌丛总生物量接近,各器官生物量的比例亦非常一致,干:叶:枝:皮的比值分别为4.09:2.03: 1.82:1.00和4.12:2.03:1.91:1.00,即属于4:2:2:1类型。
|
|
由于生境的差异,引起环境因子如温度、湿度、土壤厚度和含水量等不同,导致不同生境下川滇高山栎各器官生物量在垂直分布上产生差异,形成特定的产量结构。随着海拔升高,川滇高山栎优势单株产量降低,海拔2 720 m为14.873 kg,海拔2 920 m的干燥生境为3.392 kg,海拔3 120 m为3.147 kg。单株地上部分生物量分布格局分为2种类型,一类在海拔2 720~2 920 m处,呈纺锤形分布,单株地上部分生物量分布在树干中部1.0~3.0 m处为最多,约占总生物量的60%~70% (图 2,a, b, c),另一类在海拔3 020~3 120 m处或低海拔的干旱生境,呈金字塔形分布,个体地上部分生物量分布随株干的升高而降低,集中分布在0~1.0 m处,占总生物量的60%以上,0~2.0 m处的生物量占总生物量的94%~99% (图 2,d、e、f)。这显然与植株高度变小有关。

|
图 2 不同海拔高度或生境川滇高山栎优胜单株生物量结构(a, b, c:海拔2 720~2 920 m; d, e, f:海拔3 020~3 120 m) Fig. 2 Above ground biomass structure of Q. aquifolioides at different altitude and habitats(a, b, c: 2 720~2 920 m; d, e, f:3 020~3 120 m) |
对卧龙自然保护区不同海拔高度川滇高山栎灌丛地上部生物量的研究结果表明:
1) 用地径(D)、树高(H)估测单株林木器官生物量的适合模型为指数模型和幂函数模型,以指数模型为最佳; 而用D2H估测单株林木器官生物量的适合模型为直线模型和指数模型,以直线模型为最佳,指数模型也能达到估测效果。所筛选的统计模型相关系数均较高,为0.882~0.996,达到极显著水平,可在实践中应用。
2) 川滇高山栎灌丛平均总生物量为25.22 t·hm-2,各组分生物量为川滇高山栎>枯枝落叶层>伴生灌木层>苔藓层>草本层,分别为18.21、5.98、0.46、0.42和0.16 t·hm-2。分别占总生物量的72.20%、23.71%、1.80%、1.66%和0.63%。
3) 在海拔2 920 m以下,川滇高山栎灌丛生物量差异不大; 海拔3 020 m处下降非常明显,2个海拔高度总生物量差异达3倍以上; 海拔2 720 m处和草甸接壤处(海拔3 120 m)总生物量差异达5倍以上。
4) 川滇高山栎种群总生物量为18.21 t·hm-2,各器官生物量大小为:干>枝>叶>皮,分别为7.88、4.90、3. 61和1.83 t·hm -2,分别占种群总生物量的43.28%、26.88%、19.82%和10.02%。
5) 川滇高山栎地上部分器官生物量大小为:干>叶>枝>皮; 地上部分器官生物量的比例具有一定规律,在海拔2 720~2 920 m处,其比例约为4:3:2:1,在海拔3 020~3 120 m处,其比例约为4:2:2:1。
6) 随着海拔升高,川滇高山栎优势单株产量降低,地上部生物量分布格局分为2种类型:在海拔2 720~2 920 m处呈纺锤形分布,地上部生物量主要分布在株干中部2.0~3.0 m处,约占地上部总生物量的60%~70%;在海拔3 020~3 120 m处或低海拔的干旱生境,呈金字塔分布,地上部生物量分布随株干的升高而降低,集中分布在0~1.0 m处,占地上部总生物量的60%以上。
方精云, 刘国华, 张舒寰. 1993. 分布区西缘油松种群生长特征. 植物生态学与地植物学学报, 17(4): 305-316. DOI:10.3321/j.issn:1005-264X.1993.04.008 |
冯宗炜, 陈楚莹, 张家武, 等. 1982. 湖南会同县两种森林群落的生物生产力. 植物生态学与地植物学丛刊, 6(4): 257-267. |
冯宗炜, 王效科, 吴刚. 1999. 中国森林生态系统生物量与生产力. 北京: 科学出版社.
|
江洪, 林鸿荣. 1985. 飞播云南松林生物量与生产力的系统研究. 四川林业科技, 6(2): 6-10. |
姜汉侨. 1980. 云南植被的分布特点及地带性规律. 云南植物研究, 2(1): 23-31. |
木村允. 1981.陆地植物群落的生产量测定法(1976).姜恕, 等译.北京:科学出版社, 70-71
|
金振洲. 1979. 云南常绿阔叶林的类型及其特征. 云南植物研究, 1(1): 90-105. |
金振洲, 区普定. 1981. 我国的硬叶常绿阔叶林. 云南大学学报, 3(2): 13-20. |
金振洲. 1987.硬叶常绿阔叶林//吴征镒, 朱彦丞主编.云南植被.北京: 科学出版社
|
李文华. 1985. 西藏森林. 北京: 科学出版社.
|
刘庆. 1999. 斑苦竹无性系植物种群生态学研究. 成都: 成都科技大学出版社.
|
杨钦周. 1990. 中国喜马拉雅地区硬叶栎林的特点与分类. 植物生态学与地植物学学报, 14(3): 197-211. |
杨玉坡, 李承彪. 1992. 四川森林. 北京: 中国林业出版社.
|
徐润清, 管中天.1992.川滇高山栎林//杨玉坡, 李承彪主编.四川森林.北京:中国林业出版社, 634-645
|
中国植被编辑委员会. 1980. 中国植被. 北京: 科学出版社.
|
钟章成, 曾波. 2001. 植物种群生态研究进展. 西南师范大学学报:自然科学版, 26(2): 230-236. |
周立江, 管中天.1992.川滇高山栎灌丛//杨玉坡, 李承彪主编.四川森林.北京:中国林业出版社, 736-741
|
周浙昆. 1993. 中国栎属的地理分布. 中国科学院研究生院学报, 10(1): 95-108. |
Fujimori T, Kawanabe S, Saito H. 1976. Biomass and primary production in forests of three major vegetation zones of the Northwestern United States. Journal of Japan Forest Society, 58(10): 360-373. |
Grubler A. 1993. Enhancing carban sink. Energy, 18: 499-522. DOI:10.1016/0360-5442(93)90025-9 |
Houghton R A, Hobbie J E, Mellilo J M. 1983. Changes in the carbon content of terrestrial biota and soils between 1860 and 1980: A net release of CO2 to the atmosphere. Ecology Monograph, 53: 235-262. DOI:10.2307/1942531 |
Houghton R A, Boone R D, Mellilo J M. 1985. Net flux of carbon dioxide from tropical forests in 1980. Nature, 361: 617-620. |
Houghton R A, Boone R D, Fruc J R. 1987. The flux of carbon from terrestrial ecosystems to the atmosphere in 1980, due to changes in land use: geographic distribution of the global flux. Tellus, 39(B): 122-139. |
Huxley J S. 1931. Notes on differential growth. America Nature, 65: 289-315. DOI:10.1086/280374 |
Iverson L R, Brown S, Grainger A. 1993. Carbon sequestration in tropical Asia: an assessment potential and suitable forest lands using geographical information system analysis. Climate Research, 3: 23-38. DOI:10.3354/cr003023 |
Iverson L R, Brown S, Prasad A, et al. 1994.Use of GIS for estimating potential and actual forest biomass for continental South and Southeast Asia//Dale V H eds. Effects of land use change on atmospheric CO2 concentrations: south and southeast asia as a case study. New York: Springer_Verlag, 67-116
|
Kittredge J. 1944. Estimation of the amount of foliage of trees and shrubs. Journal of Forest, 42(11): 905-912. |
Monsi M. 1968.Mathematical models of plant communities//Eckardt F E ed. Functioning of terrestrial ecosystems at the primary production level: Proceeding Copenhagen Symposium. Natural Resources Research. Paris: UNESCO, 5: 349-358
|
Ruard G A, Martin G J, Bockheim J G. 1987. Comparison of constant and variable allometric ratios for estimating Populus tremuloides biomass. Forest Science, 33(2): 294-300. |