 2006, Vol. 42
2006, Vol. 42文章信息
- 徐华潮, 施祖华, 吴鸿.
- Xu Huachao, Shi Zuhua, Wu Hong.
- 球孢白僵菌对竹梢凸唇斑蚜的毒力
- Virulence of Beauveria bassiana against Takecallis taiwanus(Takahashi)
- 林业科学, 2006, 42(1): 85-89.
- Scientia Silvae Sinicae, 2006, 42(1): 85-89.
-
文章历史
- 收稿日期:2003-11-11
-
作者相关文章


2. 浙江大学植物保护系 杭州 310029
2. Department of Plant Protection, Zhejiang University Hangzhou 310029
竹梢凸唇斑蚜(Takecallis taiwanus)是竹子的主要害虫之一,寄主多,分布广泛,为害严重(张广学等,1983;胡国良等,2001)。若蚜、成蚜均有红、绿2种颜色,成蚜均为有翅蚜(徐华潮等,2003)。目前对它的防治主要依靠化学农药,其他防治方法应用很少,从而带来了一系列的负面影响(胡国良等,2001)。对该蚜虫开展综合治理,特别是生物防治方法研究有着广阔的应用前景。然而目前尚未见该领域系统的研究工作。笔者研究了球孢白僵菌(Beauveria bassiana)对竹梢凸唇斑蚜的致病力,旨在为该蚜虫的生物防治提供科学依据。
1 材料与方法 1.1 供试菌株球孢白僵菌菌株F-263是从日本松墨天牛(Monochamus alternatus)幼虫上分离获得;Zhe-B由浙江临海白僵菌厂提供;REBb01是从安徽木麻黄天牛(Diastocera wallichi)幼虫上分离获得;B12是从丽水马尾松毛虫(Dendrolimus punctatus)幼虫上分离获得;Bxs是从浙江象山松墨天牛幼虫上分离获得。菌株供试前经侵染竹梢凸唇斑蚜成蚜复壮纯化后暂时保存在温度为4~6 ℃的冰箱中。本试验用菌株为在含有PPDA培养基上在光照培养箱内(温度25 ℃±1 ℃,相对湿度80%,光照14L:10D)培养1周后获得的分生孢子粉。
1.2 供试虫源从未施农药的雷竹(Phyllostachys praecox)林中采回健康的竹梢凸唇斑蚜成蚜做虫源,仿照叶子圆片法(刘树生,1987)以雷竹叶片为饲料连续饲养(温度25 ℃±1 ℃,相对湿度80%,光照12L:12D),建立室内饲养群。生物测定时所用蚜虫为室内饲养群中同一批若蚜在相同条件下饲养,羽化后1~2 d内的成蚜。
1.3 生物测定用含有0.1%吐温-80润湿剂的KH2PO4缓冲液(0.003 mol·L-1)将各菌株的分生孢子粉配制成系列梯度悬浮液,从低到高依次为1×104、1×105、2.5×105、5.0×105、7.5×105、1×106、1×107个孢子·mL-1。以不含分生孢子的KH2PO4缓冲液(0.003 mol·L-1)作为对照。各浓度及对照供试蚜虫数为30头。采用浸渍法接菌(刘银泉等,1999;Feng et al., 1990)。接菌后的蚜虫用湿润毛笔轻轻移到小号养蚜塑杯(上口径2.6 cm,下口径2.4 cm,高4.3 cm)中,仿叶子圆片法在培养箱内(温度25 ℃,相对湿度80%~90%,光照12L:12D)单头饲养。连续10 d每天定时观察、记录新产的若蚜数和成蚜的死亡情况,将蚜尸及时移出依次编号保湿培养,2周后镜检,以蚜尸上有球孢白僵菌长出作为有效致死标准。
1.4 数据分析采用时间-剂量-死亡率模型(time-dose-mortality model, TDM),又称互补重对数模型(complementary log-log model, CLL),对试验数据进行模拟和各参数估计。具体可参考冯明光等(1996;1997)所报道的方法,整个运算过程用DPS计算机软件(唐启义等,2002)完成。
2 结果与分析竹梢凸唇斑蚜接种球孢白僵菌后逐日存活数逐渐减少,即累积死亡率逐渐上升。而对照组在10 d内未见有死亡。可见供试5个菌株对竹梢凸唇斑蚜都表现出一定的致病性,但毒力强弱因菌株而有所不同。同一菌株处理下,竹梢凸唇斑蚜的累积死亡率随孢子浓度增加而上升。接种后的蚜虫存活数逐日减少,说明累积死亡率是随着处理后时间的延长而递升,但上升幅度及高峰值的出现时期则因菌株和浓度而异。在高剂量下(1×107个孢子·mL-1),菌株B12和Bxs的致死率可高达100%,而菌株F-263和Zhe-B的致死率只有92.59%;尽管各菌株均在第2—3天即出现染病而死的蚜虫,至第7—8天达到死亡高峰,但期间表现出的曲线上升幅度不同,其中B12、Bxs菌株上升幅度最大,在第7天累积死亡率已分别高达100%和96.4%,其次为菌株Zhe-B和REBb01,第8天的累积死亡率分别达92.59%和89. 65%,最慢的是F-263菌株,到第8天累积死亡率仅为85.19%。菌株B12、Bxs在高浓度下出现较高死亡率的时间明显早于其他菌株,显示出这2个菌株对该蚜虫有较强的毒力。中浓度处理(1×105~1×106个孢子·mL-1)后,死亡大都始于第3天或第4天,经过5 d左右达到高峰并渐趋平缓。低浓度处理(1×104~1×105个孢子·mL- 1)后,蚜虫开始出现死亡时间更加延迟,各菌株间也有明显差异,菌株B12处理后在第3天即可出现死亡,菌株Zhe-B处理后直至第6天才观察到蚜虫死亡,且死亡率在第8天就已基本稳定,而菌株Bxs、REBb01处理后,蚜虫死亡在第4、5天开始出现后一直可持续到第10天。这些结果表明:累积死亡率不仅是时间相关函数,还是剂量相关函数,这也说明综合考虑时间和剂量效应的重要性。
用DPS数据处理系统软件对实验数据进行时间-剂量-死亡率模型的拟合,获得条件死亡率模型的剂量效应参数(β)和时间效应参数(γj)的估计值,继而估计出累积死亡率模型的时间效应参数(τj)列于表 1(下标j表示处理后第j天)。条件死亡率模型中各参数估计值的t测验均达到极显著水平(P<0.000 1)。条件死亡概率的时间效应参数γj在接种后第2天至第7天或第8天呈递增趋势,随后呈递减趋势。这与试验观察中的死亡高峰期相吻合,表明几种供试菌株对竹梢凸唇斑蚜的杀蚜潜伏期约为7~8 d。
|
|
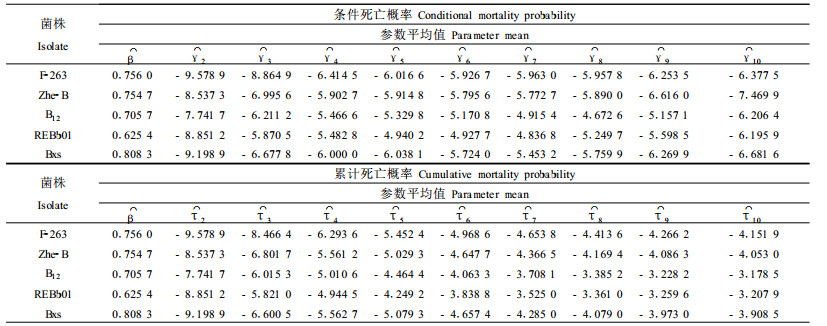
在表 1中,5个菌株的参数β值介于0.625 4~0.803 8之间,说明所试菌株虽然对竹梢凸唇斑蚜都有致死效果,但剂量效应因菌株而异,该斜率值范围明显小于国外报道的多种化学杀虫剂作用于昆虫的同一参数的估计范围(1.2~4.5)(Preisler et al., 1989)。显示所测菌株的杀蚜效果比化学杀虫剂要缓慢,这符合一般微生物制剂的作用特点。实际上,真菌杀虫剂对昆虫寄主的侵染除了与时间及剂量相关外,还受环境条件的影响,其中温度是重要的影响因子。温度不仅影响杀虫真菌孢子的萌发和侵染速率(Feng et al., 1994),还影响毒力的表达(刘银泉等,2000)。
据Hosmer-Lemeshow检验,模型拟合无显著异质性存在(表 2),说明CLL模型适于此类数据的分析。在成功拟合各个参数后,可用于剂量与时间效应的估计及二者间互作效应的评价。
|
|

根据表 1中有关累计死亡率模型的参数模拟值,利用CLL模型可分别估计出不同菌株对竹梢凸唇斑蚜在接种后不同时间的剂量效应值及其标准误。竹梢凸唇斑蚜在不同菌株下随时间变化的lg(LC50)和lg(LC90)及其95%置信限见图 1。由图 1可以看出各菌株的lg(LC50)和lg(LC90)在接种处理后前7 d内变化很大,7 d后曲线坡度趋向平缓,表明死亡率在前期变化较大,后期变化较小。因此根据浓度效应比较各菌株毒力相对强弱时最好选择在接种处理7 d后的某1天。从图 1还可看出,致死剂量是时间的函数,接菌后随着时间的延续,lg(LC50)和lg(LC90)渐次减少,表明剂量效应逐渐增强,同样,随剂量的增加,时间效应也相应增强,这符合杀虫微生物的一般作用规律。

|
图 1 随时间变化的球孢白僵菌对竹梢凸唇斑蚜的lg(LC50)和lg(LC90)及其95%置信限 Fig. 1 Time-dependent lg(LC50), lg(LC90) and their 95% confidence intervals of Beauveria bassiana against Takecallis taiwanus |
供试菌株不同浓度对竹梢凸唇斑蚜致死率的时间效应也可由CLL模型估算出,除最终累积死亡率低于50%的低剂量处理外,其他剂量处理的致死中时LT50均能从累积死亡概率模型用插值法作出估计。图 2为各菌株不同浓度下的LT50。从图中可见,时间效应与剂量呈负相关趋势,即LT50值随浓度增加而减少。值得注意的是,在组建CLL模型的有效时区(上限为试验的最后观察日数)内,能产生50%死亡概率的最低浓度在不同菌株间差异较大,该浓度值越小,表示该菌株对竹梢凸唇斑蚜的毒力越强,由此可基本识别出毒力最强的菌株为B12,最弱的为F-263。在同一浓度下,LT50值越小,该菌株的毒力相对较强,如在1×106个孢子·mL-1浓度下,5个菌株的LT50值顺序为:B12(3.63 d)<Bxs(3.70 d)<REBb01(4.31 d)<Zhe-B(4.34 d)<F-263(5.20 d),说明以上各菌株毒力渐次变弱,此结果与浓度效应所得结论一致。

|
图 2 随剂量而变化的球孢白僵菌对竹梢凸唇斑蚜的LT50 Fig. 2 Dose-dependent LT50 of B. bassiana against T. taiwanus |
根据所估计参数而建立的CLL模型,可以完整地描述所测球孢白僵菌菌株对竹梢凸唇斑蚜的时间与剂量效应,即随时间和剂量而变化的死亡率,如图 3所示。在实验涵盖的时间与剂量范围内,模拟的死亡概率是一个曲面。根据曲面的高度和变化趋势,可以看出不同菌株对竹梢凸唇斑蚜的毒力强弱差异。从剂量与时间的综合效应考虑,菌株B12、Bxs对竹梢凸唇斑蚜的毒力最强,其次是REBb01和Zhe-B,菌株F-263的毒力相对较弱。

|
图 3 球孢白僵菌作用下竹梢凸唇斑蚜随时间和浓度而变化的累积死亡率 Fig. 3 Cumulative mortality of T. taiwanus attributed to inoculation of B. bassiana at varying time and dose |
研究结果表明:来自不同虫源的5个球孢白僵菌菌株对竹梢凸唇斑蚜都表现出一定的致病力。总体而言,菌株B12和菌株Bxs表现出较强毒力,但与化学杀虫剂相比,其杀虫效果均较为缓慢。尽管球孢白僵菌对竹梢凸唇斑蚜的致病力以及接种后对该蚜虫种群增殖的控制都存在一定的局限性,但不能因此而否定了球孢白僵菌作为竹梢凸唇斑蚜微生物防治因子的潜力。本研究中所用菌株由于保存时间较长,使用时多次转接可能也是引起其侵染力下降的一个因素。考虑到球孢白僵菌自身生长所需条件及环境因子的影响,若单独使用球孢白僵菌可能很难对竹梢凸唇斑蚜的种群增长有很大的控制作用,有必要将它作为该蚜虫综合防治中一项措施继续加以研究和利用。
冯明光. 1997. 迁徙蚱蜢经口器摄入感染球孢白僵菌的时间与剂量效应. 浙江农业大学学报, 23(5): 491-498. |
冯明光, 唐启义, 胡国成, 等. 1996. 球孢白僵菌对七种蚜虫的感染反应—时间-剂量-死亡率模型分析. 应用基础与工程科学学报, 4(1): 22-33. |
胡国良, 俞彩珠, 楼君芳, 等. 2001. 竹梢凸唇斑蚜的生物学特性与防治. 浙江林学院学报, 18(3): 294-296. DOI:10.3969/j.issn.2095-0756.2001.03.017 |
刘树生. 1987. 介绍一种饲养蚜虫的方法——新的叶子圆片法. 昆虫知识, 24(2): 113-115. |
刘银泉, 冯明光, 刘树生, 等. 2000. 不同温度下球孢白僵菌对桃蚜的毒力. 中国生物防治, 16(2): 56-60. |
刘银泉, 冯明光, 刘树生, 等. 1999. 球孢白僵菌对桃蚜的毒力测定. 植物保护学报, 26(4): 347-352. DOI:10.3321/j.issn:0577-7518.1999.04.013 |
唐启义, 冯明光. 2002. 实用统计分析及其DPS数据处理系统. 北京: 科学出版社, 188-195.
|
徐华潮, 施祖华. 2003. 温度对竹梢凸唇斑蚜种群增长的影响. 浙江大学学报:农业与生命科学版, 29(4): 439-443. |
张广学, 钟铁森. 1983. 中国经济昆虫志:同翅目·蚜虫类(一). 北京: 科学出版社, 157-158.
|
Feng M G, Johnson J B. 1990. Relative virulence of six isolates of Beauveria bassiana on Diuraphis noxia (Homoptera: Aphididae). Environmental Entomology, 19(3): 785-790. DOI:10.1093/ee/19.3.785 |
Feng M G, Poprawski T J, Khachatourians G G. 1994. Production, formulation and application of the entomopathogenic fungus, Beauveria bassiana for insect control: current status. Biocontrol Science and Technology, 4: 3-34. DOI:10.1080/09583159409355309 |
Preisler H K, Robertson J L. 1989. Analysis of time-dose-mortality data. Journal Economic Entomology, 82(6): 1 534-1 542. DOI:10.1093/jee/82.6.1534 |