 2006, Vol. 42
2006, Vol. 42文章信息
- 张含国.
- Zhang Hanguo.
- 落叶松F2代杂种优势的稳定性研究
- Study on Heterosis Stability of F2 Generation of Hybrid Larch
- 林业科学, 2006, 42(1): 49-54.
- Scientia Silvae Sinicae, 2006, 42(1): 49-54.
-
文章历史
- 收稿日期:2004-07-05
-
作者相关文章


杂种优势在生物界普遍存在,那么如何利用杂种优势就显得非常重要。通常情况下,获得经过测定的优良杂种后,采用无性繁殖,然后大规模造林,从而提高木材产量和质量。落叶松属难生根树种,在高寒地区生根率低且不稳定,所以无性繁殖很难大规模推广,因此,杂种的有性利用显得非常重要。根据性状遗传的基本规律,F2代群体内必然出现性状的分离和重组,F2与F1代相比,杂种优势的表现显著地下降。F2代能否利用还存在不同的看法,巴西起源的巨桉(Eucalyptus grandis)是一个明显退化的例子, 杂种F2代出现5%~10%不需要的类型(Brune et al., 1981);Hyun等(1976)发现刚松×火炬松(Pinus rigida×P.taeda)的子代在7年生时F2与F1代高度相近,F2及F3代可以利用。南京林业大学获得的鹅掌楸×北美鹅掌楸(Liriodendron chinense×L.tulipifera)杂种,其F2与F1代基本相似,可用于生产繁殖(王明庥,2001)。
落叶松种间杂种F1代生长初期具有一定的杂种优势,不同组合杂种优势不同。日本落叶松×兴安落叶松(Larix kaempferi×L.gmelinii)F1代高生长超过亲本平均数的87.5%,日本落叶松×长白落叶松(L.kaempferi×L.olgensis)苗期生长量为母本的154%,兴安落叶松×长白落叶松苗期生长量为母本的107%(潘本立等,1981;王景章等,1980;杨书文等,1985;张含国等,1998)。11年生欧洲落叶松×日本落叶松(L. decidua×L.kaempferi)的树高、胸径和材积分别超过欧洲落叶松自由授粉子代的5%~22%、7%~31%、29%~122%,最优家系树高、胸径和材积分别超过欧洲落叶松的5%~33%、27%~50%、85%~170%(杨书文等,1994)。那么落叶松F2代生长优势如何?李希才等(1995)证明落叶松杂种二代也具有显著的增产能力,7年生时各家系树高分别比落叶松杂种种子园、长白落叶松种子园和当地一般生产对照高10%~14.9%、22.7%~39.6%、32.4%~50.7%。本文数据源于1988年营建的2组测定林,主要探讨杂种落叶松F1代与F2代是否存在严重的分离现象,生长后期杂种优势如何,从而更好地确定杂种落叶松的有效利用等问题。
1 材料与方法 1.1 材料1965年在黑龙江省林口县青山林场营建日本落叶松、兴安落叶松无性系种子园,其中日本落叶松优树选自当地日本落叶松人工母树林,兴安落叶松优树来自于小兴安岭汤汪河林业局东汤林场。1977年在种子园中利用套袋控制授粉技术进行杂交制种(长白落叶松优树花粉由当地人工母树林采集),1977年秋采种,1978年分家系播种育苗,1980年营建F1代杂种实生种子园,共有7个F1代杂种家系(日3×兴2、日3×兴9、日12×兴9、日5×兴9、日5×长77-3、日5×长78-3、兴7×日77-2),单行20株为一个家系。1988年利用采自F1代杂种实生种子园种子,营建2组子代测定林。本文分析数据来源于子代测定林1994、1995、1996、2002年秋每木调查数据。
第1组试验材料1988年在F1代实生种子园中分家系采种、调制,1989年播种育苗(F2代),1991年上山定植(9116区),小区20株单行排列,重复4次,处理数10个(7个处理为杂种种子园家系),其中包含3个对照:长白落叶松种子园混合种子(CK1)、小北湖优良种源种子(CK2)、当地生产种子(CK3)。
第2组试验材料1988年在实生种子园中对F1代日5×兴9的15个单株采集自由授粉种子、调制,1989年播种育苗(F2代),1991年按随机区组设计定植(9101区),20个处理[1~15为日5×兴9各家系自由授粉获得的F2代,16为日5×兴9混,17为日本落叶松5号(日5),18为兴安落叶松9号(兴9),19为长白落叶松种子园混(CK1),20为长白落叶松小北湖优良种源(CK2)],单行小区20株,重复4次。
1.2 统计分析方法杂种优势、单株遗传力、家系遗传力以及遗传增益计算参见王明庥等(2001)、乔纳森(1981)和佐贝尔等(1990):杂种优势(%)=(F1平均值-较好亲本平均值)/双亲平均值×100;超亲优势(%)=(F1平均值-较好亲本平均值)/较好亲本平均值×100。hi2=4σf2/(σf2+σfb2+σe2);
hf2=σf2/(σf2+σfb2/b+σe2/nb)。式中:σf2、σfb2、σe2分别为家系、区组×家系、环境方差; hf2, hi2分别为家系、单株遗传力; n为小区株数,b为区组数(重复数)。遗传增益: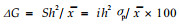
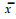
对数据进行方差分析、LSR检验等,分析采用SPSS12.0的General Linear Model中的Univariate软件(刘先勇等,2002)。分析过程中对奇异数值进行了剔除,树高、胸径和材积株数不完全一致。立木材积按平均实验形数法计算,公式为:V=(h+3)g1.3f3,式中,落叶松平均实验形数f3为0.41,所以,V=0.32(h+3)d2/10 000,式中, h为树高,d为胸径。
2 结果与分析 2.1 落叶松杂种F2代家系内的遗传分离 2.1.1 不同落叶松杂种F2代家系内遗传分离第1组试验材料主要是探讨7个杂种家系F2代的遗传分离情况(表 1)。从9116组F2代7个杂种家系内14年生高生长变异情况来看,家系内存在一定的遗传分离,但分离不大,都在平均值1倍标准差范围内。家系间变异系数(CV)的变化幅度不大,其家系CV为12.1%~21.2%,平均为17.6%,其中,日5×兴9、兴7×日77-2家系内分离稍大,日3×兴9家系内分离较小。
|
|
从12年生5个F1代杂种家系内(日3×兴2、日3×兴9、日12×兴9、日5×兴9、日5×长77-3)高生长变异情况来看,家系内也存在一定的遗传分离,除日5×长77-3家系内CV稍低外,其余几个家系相差无几,CV为9.3%~12.9%,平均为11.5%。相同5个家系F2代的CV平均为17.3%,比F1代只增加5.8%,这就是说F2代比F1代家系内变异只大5.8%。
7个杂种家系F2代6、7、8、14年生高生长的CV平均值分别为24.3%、23.9%、22.1%、17.6%,可以看出,随树龄增加,家系内分离幅度呈减小的趋势,变异渐趋稳定,14年生高生长的CV比6年生时小6.3%。
7个F2代家系内胸径生长也存在一定的遗传分离,CV为16.7%~29.2%,平均22.7%,家系间CV变幅较高生长稍大。其中,日12×兴9、兴7×日77-2家系内变异较大,分别为29.2%、25.9%;日3×兴9、日3×兴2两个家系内变异较小,分别为16.7%、19.2%。12年生的5个F1代家系内胸径生长的CV为18.1%~22.5%,平均19.9%。相同5个家系F2代的CV平均为22.0%,比F1代增加1.1%,F2代与F1代相比家系内分离不大。
F2代家系内材积生长存在的遗传分离稍大,CV为38.3%~60.4%,平均49.5%,家系间CV变幅也较大。
2.1.2 日5×兴9 F2代家系内遗传分离第2组试验材料是探讨F1代1个杂种家系日5×兴9 F2代遗传分离情况(图 1a)。数据表明15个14年生的日5×兴9 F2代家系内高生长存在一定的遗传分离,不同家系CV为11.4%~18.3%,平均13.9%,较第1组试验材料日5×兴9 F2代CV还小7.3%。家系间CV变幅也不大,其中,13、15、1和5号家系内变异稍大,12、4、11和2号家系内变异较小。从12年生日5×兴9 F1代高生长变异情况来看,家系内存在一定的遗传分离,CV为11.6%,同F2代比低2.3%,说明F2代与F1代相比家系内分离不大。6、7、8、14年生高生长的CV分别为24.5%、22.0%、17.9%、13.9%,从CV看出,日5×兴9杂种家系F2代随树龄增加,家系内遗传分离幅度也呈减小趋势,14年生高生长CV比6年生时小10.6%。

|
图 1 日5×兴9各家系树高、胸径生长分布图 Fig. 1 The distribution of height growth and diameter growth of L. kaempferi 5×L.gmelinii 9 families |
15个日5×兴9 F2代家系内胸径生长也存在遗传分离,CV为13.6%~26.8%,平均21.6%,家系间变异幅度较高生长稍大。其中,13、1、6和15号家系内变异较大,7、8、4和1号家系内变异较小(图 1b)。
家系内材积生长存在的遗传分离稍大,CV为36.8%~53.8%,平均为45.7%,家系间变异幅度也相对较大。其中,3、6、13和1号家系内变异较大,8、2、11和5号家系内变异较小。
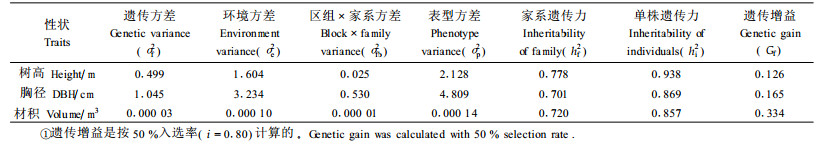
2.2 F2代杂种优势 2.2.1 7个落叶松杂种F2代遗传增益为了更准确地计算遗传力,将缺少试验数据的重复删掉,每个小区随机抽取4株树的数据,试验设计变成3个重复,小区株数为4株的随机区组排列。7个杂种落叶松F2代家系间方差分析结果表明,F2代家系间14年生树高、胸径和材积差异都达极显著水平。根据方差分量计算遗传力及遗传增益(表 2),树高、胸径、材积遗传力较高,属于强度遗传,树高、胸径的遗传增益都在10%以上,材积的遗传增益高达33%,所以F2代仍可以利用。
|
|
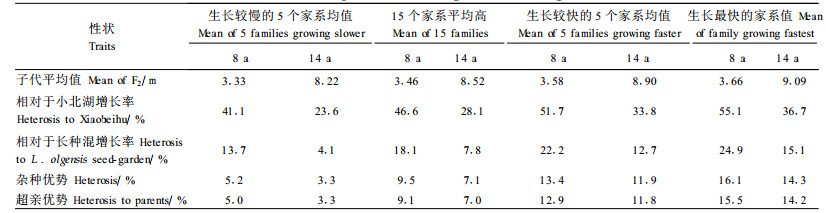
14年生20个处理试验林方差分析表明,树高、胸径和材积处理间差异极显著。树高多重检验表明,小北湖高生长最差,兴9、长种混、日5排在倒数2~4位,日5×兴9混居第9位,其余15个家系都是日5×兴9的F2代家系(图 2a)。15个F2代家系高生长平均值为8.52 m,超过小北湖28.1%,超过长种混7.8%,杂种优势为7.1%。其中,生长最快的8号家系高生长为9.09 m,超过小北湖36.7%,超过长种混15.1%;生长较快的5个家系(8、2、7、1和6号)高生长均值为8.90 m,超过小北湖33.8%,超过长种混12.7%,杂种优势为11.9%;生长较慢的5个家系高生长均值为8.22 m,超过小北湖23.6%,超过长种混4.1%,杂种优势为3.3%;生长最慢的13号家系树高超过小北湖19.8%,超过长种混0.9%。

|
图 2 日5×兴9F2代家系树高、材积生长分布图 Fig. 2 The distribution of height and volume of L. kaempferi 5×L.gmelinii 9 families |
材积LSR检验结果表明,小北湖生长最差,长种混、日5、兴9材积排在倒数2、4、7位,日5×兴9混居第9位,其余都是日5×兴9的F2代家系(图 2b)。15个F2代家系材积生长平均值为0.030 03 m3,超过小北湖123.8%,超过长种混33.8%,超过父本11.8%,超过母本17.7%,杂种优势为12.1%。其中,生长最快的8号家系材积为0.038 17 m3,超过小北湖184.4%,超过长种混70.1%,超过父本42.2%,超过母本49.6%;生长较快的5个家系材积均值为0.034 05 m3, 超过小北湖153.7%,超过长种混51.7%,超过父本26.8%,超过母本33.5%,杂种优势为27.5%;生长较慢的5个家系材积均值为0.026 38 m3, 超过小北湖96.6%,超过长种混17.6%;生长最慢的13号家系材积超过小北湖84.1%,超过长种混10.1%。
胸径生长LSR检验结果表明,小北湖生长最差,长种混、日5、兴9排在倒数2、5、8位,日5×兴9混居第8位,其余都是日5×兴9的F2代家系。15个家系F2代胸径生长超过小北湖43.8%,超过长种混15.7%,超过父本4.1%,超过母本7.3%,杂种优势为4.1%。其中,生长最快的8号家系胸径生长超过小北湖56.9%,超过长种混26.2%,超过父本13.5%,超过母本17.0%,杂种优势为13.7%;生长较快的5个家系胸径生长超过小北湖52.1%,超过长种混22.3%,超过父本10.0%,超过母本13.4%,杂种优势为10.2%;生长较慢的5个家系胸径生长超过小北湖37.2%,超过长种混10.4%,超过母本2.3%;生长最慢的13号家系胸径超过小北湖29.1%,超过长种混3.9%。
2.3 杂种优势的年龄效应随着树木年龄的增长,杂种比其亲本是否有持久的杂种优势,到目前为止,仍然没有明确的答案,需要通过适当的试验设计并进行长期的观察,才能得到确定的答案。F2代杂种落叶松的杂种优势同对照相比增产效果还是十分明显的。虽然14年生的增产效果同8年生时相比有增幅减缓趋势,但仍具有较大的杂种优势(表 3)。以小北湖种源作参照,15个家系平均值14年生高生长比8年生下降了18.5%,相比于长白落叶松种子园则下降了10.3%,杂种优势增幅下降了2.4%。以小北湖种源作参照,生长较快的5个家系平均值14年生高生长比8年生下降了17.9%,相比于长白落叶松种子园则下降了9.5%,杂种优势增幅下降了1.5%。还以小北湖种源作参照,生长量较差的5个家系平均值14年生高生长比8年生下降了17.5%,相比于长白落叶松种子园则下降了9.6%,杂种优势增幅下降了1.7%。
|
|
杂种优势的稳定性主要体现在2个方面:遗传分离程度、不同年龄阶段的杂种优势。А.С.Яблков于1967年报道日本落叶松×西伯利亚落叶松(L. kaempferi ×L. sibirica)F2代的生长优于亲本西伯利亚落叶松,但由于基因重组子代个体间分化较大,回交种要比杂种自由授粉子代生长更为整齐,提倡杂种落叶松的越代利用(杨书文,1994)。尽管不同学者在评价杂种优势时所用的材料和方式不同,但F2代都表现出超出其纯种亲本的杂种优势,但都比F1代杂种优势要低,并且与F1代相比变异稍大(仓桥昭夫,1988)。
本文结果表明,7个F2代杂种家系内14年生高生长存在一定的遗传分离,在平均值1倍标准差范围内。家系间变异系数(CV)的变化幅度不大,其家系CV为12.1%~21.2%,平均17.6%。相同5个家系F2代的CV平均为17.3%,比F1代CV只增加5.8%,这就是说F2代比F1代家系内变异只大5.8%。12年生F1代日5×兴9高生长CV为11.6%,同F2代比低2.3%,说明F2代与F1代相比家系内分离不大。由这2组材料来看,F2代家系内的遗传分离并不比F1代有明显的增加,F2代还是可以利用的。7个杂种家系F2代6、7、8、14年生高生长的CV平均值分别为24.3%、23.9%、22.1%、17.6%,14年生高生长的CV比6年生时小6.3%;日5×兴9的F2代树高、胸径和材积家系内也存在一定的变异,14、8、7和6年生树高CV也呈由小到大的趋势(13.9%、17.9%、22.0%和24.5%),这说明杂种F2代的生长初期分离较大,这与环境作用有关。随着树龄增加,家系内遗传分离幅度呈减小的趋势,也就是家系内遗传分离将会趋于稳定状态。
杂种的早期优势是否能一直保持下去而形成真正的杂种优势,或者这只是暂时的一个早期速生阶段?一直没有一个明确的答案。据Rcek等于1977年报道,欧洲落叶松×日本落叶松的F2和F3代生长仍然超过欧洲落叶松,因此,在欧洲落叶松×日本落叶松杂种的F1代中选择优良单株营建种子园生产F2代种子(杨书文等,1994)。Gothe等(1980)分析了欧洲落叶松×日本落叶松阶段性的生长增益。Gothe(1987)在德国的试验表明,欧洲落叶松×日本落叶松从树龄20年开始杂种在绝对产量优势上表现出微小的下降,而与亲本子代相比的相对产量优势也有明显下降。Keiding(1980)和Reck(1980)的试验结果证明了同样的结论,欧洲落叶松×日本落叶松杂种在最初10年表现出最大的杂种优势。Scamoni(1977)也支持这个观点。但从一个法国试验中获得的数据(Ferrand et al., 1985)与这些结果不一致:26年的欧洲落叶松×日本落叶松杂种不仅保持其绝对优势,相对其亲本也保持着材积上的相对优势。Rohmeder等(1959)发现日本落叶松×欧洲落叶松的F2代和回交种有生长优势,但比F1代的杂种优势要弱。
F2代杂种落叶松的杂种优势以及同对照相比增产效果还是十分明显的。7个杂种F2代家系树高、胸径、材积遗传力较高,属强度遗传,遗传增益都在10%以上,材积遗传增益高达33%,所以F2代仍可利用。本文14年生日5×兴9 F2代优势明显,树高、胸径、材积分别超过优良种源小北湖28.1%、43.8%、123.8%,超过长种混7.8%、15.7%、33.8%,杂种优势为7.1%、4.1%、12.1%。虽然14年生的增产效果同8年生相比增幅减缓,但仍具有较大的杂种优势。以小北湖种源作参照,15个家系14年生高生长平均值比8年生下降了18.5%,相比于长白落叶松种子园则下降了10.3%,杂种优势增幅下降了2.4%。以小北湖种源作参照,生长较快的5个家系14年生高生长平均值比8年生下降了17.9%,相比于长白落叶松种子园则下降了9.5%,杂种优势增幅下降了1.5%。以小北湖种源作参照,生长较差的5个家系14年生高生长平均值比8年生下降了17.5%,相比于长白落叶松种子园则下降了9.6%,杂种优势增幅下降了1.7%。杂种优势下降的原因之一是杂种落叶松生长快,需要的营养多,而环境营养缺乏,满足不了正常生长所需的营养物质。
F2代确实存在分离现象,但分离不是很大,而且随树龄增加,分离渐趋稳定,并具有一定的杂种优势,再加上落叶松从造林到主伐需经过几次间伐,因此,主伐时的生长量还是较高的。通过选择,利用最优良的杂种F1代优树组建杂种无性系种子园是目前我国高寒地区大量生产遗传增益较高的杂种种子途径之一。
仓桥昭夫. 1988. 落叶松杂交育种的研究. 东京大学演习林报告, 79: 1-94. |
李希才, 姚君, 王维臣, 等. 1995. 杂种二代落叶松增产能力的研究. 林业科技通讯, (2): 2-22. |
刘先勇, 袁长迎, 段宝福, 等. 2002. SPSS 10.0统计分析软件与应用. 北京: 国防工业出版社.
|
潘本立, 艾正明, 韩承伟, 等. 1981. 落叶松立木杂交方法及育种优势的研究. 林业科学, 17(3): 325-330. |
乔纳森W赖特. 1981.森林遗传学.郭锡昌, 胡承海, 译.北京: 中国林业出版社, 288-295
|
王景章, 丛培艳. 1980. 落叶松杂交育种及F1性状遗传. 林业科学, 16(1): 49-52. |
王明庥. 2001. 林木遗传育种学. 北京: 中国林业出版社.
|
杨书文, 鞠永贵, 张世英, 等. 1985. 落叶松杂种优势的研究. 东北林业大学学报, 13(1): 30-36. |
杨书文, 王秋玉, 夏得安. 1994. 落叶松的遗传改良. 哈尔滨: 东北林业大学出版社.
|
张含国, 袁桂华, 李希才, 等. 1998. 杂种落叶松生长和材质联合选择的研究. 东北林业大学学报, 26(3): 12-18. |
佐贝尔B J, 约翰·塔尔伯特.1990.实用林木改良.王章荣, 陈天华, 等译.哈尔滨: 东北林业大学出版社, 407-451
|
Brune A, Zobei B J. 1981. Genetic base populations, gene pools and breeding populations for Eucalyptus in Brazil. Science, 102(2641): 135-142. |
Ferrand J C, Bastien J C. 1985. Bilan à 26 ans d'une plantation comparative de mélèzes. Rev For Fr, 37(6): 441-448. |
Gothe H. 1987. Ein Kreuzungsversuch mit Larix europaea D.C., Herkunft Schlitz, und Larix leptolepis Gord. Allg Forstztg, 158(1): 1-3. |
Gothe H, Schober R, Bork H. 1980. Ein Kreuzungsversuch mit Larix europaea D.C., Herkunft Schlitz, und Larix leptolepis Gord. Allg Forstztg, 151(6-7): 101-112. |
Hyun S K. 1976. Interspecific hybridization in Pines with the special reference to Pinus rigida×taeda. Silvae Genetica, 25(5-6): 188-191. |
Keiding H. 1980. Hybrid laerkens vaekst og tilpasningseven Ⅰ forhold til de rene arter. Dansk Skovforen Tidsskr, 65: 204-234. |
Reck S. 1980. Untersuchung über das Holz der Hybridlärche. Allg Forstztg, 151(6-7): 117-120. |
Scamoni A. 1977. Uber lärchenhybriden aus freier Bestaubung und über die weitere Entwicklung gelenkter Lärchenkreuzungen. Beitr Forstwirtsch, 11(1): 21-26. |
Rohmeder E, Schönbach H.1959.Genetik und Züchtung der Waldbaume. Parey, Berlin
|