 2006, Vol. 42
2006, Vol. 42文章信息
- 谭晓风, 胡芳名, 谢禄山, 石明旺, 张党权, 乌云塔娜.
- Tan Xiaofeng, Hu Fangming, Xie Lushan, Shi Mingwang, Zhang Dangquan, Wuyun Tana.
- 油茶种子EST文库构建及主要表达基因的分析
- Construction of EST Library and Analysis of Main Expressed Genes of Camellia oleifera Seeds
- 林业科学, 2006, 42(1): 43-48.
- Scientia Silvae Sinicae, 2006, 42(1): 43-48.
-
文章历史
- 收稿日期:2004-12-09
-
作者相关文章


2. 中南林业科技大学资源与环境学院 长沙 410004
2. College of Resources and Environment, Central-South Forestry University Changsha 410004
油茶(Camellia oleifera)是我国长江流域及以南各省区的重要木本油料树种(中国科学院植物研究所,1971),种仁含油率约为55%。所产茶油富含90%的油酸、亚油酸等不饱和脂肪酸,是易消化耐贮藏的优质食用植物油(何方等,2004)。目前国内外对油茶的研究集中在选优栽培方面,分子水平的研究集中在种质资源的RAPD分析(陈永忠等, 2005; 黄永芳等, 2005; 2004;张云等, 2003),而关于种子油脂转化的基因研究尚未见报道。本文以生产上广泛应用的2个油茶优良无性系湘林1号和湘林4号的近成熟种子混合样品为材料,构建了cDNA文库(胡芳名等, 2004),并对油茶种子在油脂转化高峰期有关基因的表达进行了鉴定,构建了EST文库,为开发油茶优质基因资源,分离克隆相关基因及油脂植物的遗传改良奠定了物质和技术基础。
1 材料与方法 1.1 材料于2003年9月中旬,从湖南省林业科学研究院油茶采穗圃采集正处油脂转化高峰期的油茶优良无性系湘林1号和湘林4号的近成熟种子。
1.2 方法 1.2.1 油茶种子的mRNA提取总RNA提取参照Chang等(1993)的方法; mRNA的分离纯化采用磁珠法。
1.2.2 cDNA文库构建mRNA分离纯化后,在SuperScript Ⅱ-RT反转录酶和其他酶的作用下合成第一链cDNA和第二链cDNA; 经双链末端补平、加接头、酶切后,回收cDNA; 然后将cDNA片段与载体pBluescript Ⅱ连接; 最后将重组cDNA转入大肠杆菌(Escherichia coli)菌株DH10B中,构建成油茶种子cDNA文库(骆蒙等,2001; 钟华等,2003; 裴凌鹏等,2003; 张光祥等,2003; 刘长征等,1995; 江昌俊,1995)。
细菌培养和质粒DNA抽提参考《分子克隆实验指南》(萨姆布鲁克等,1999)的方法进行。随机挑取2 327个克隆,委托北京华大公司进行3′端单向序列测定。
1.2.3 生物信息学分析采用NCBI的BLAST软件对所测核苷酸序列与GenBank中的非冗余数据库(non-redundant database,NR)比对。这一数据库是去除GenBank、EMBL和DDBJ中所有相同核酸序列整合后获得的最为全面的已知基因的数据库, 其中还包括部分基因组的序列, 同时参照EST、STS、GSS等数据库。
2 结果与分析 2.1 油茶cDNA文库构建以油茶优良无性系近成熟种子成功地构建了cDNA文库(胡芳名等,2004)。经检测,其库容量高达1.8×106,重组率为98.1%,插入片段的平均长度约为1 100 bp,表明构建的油茶种子cDNA文库具有较高的质量,能够满足后续的EST文库构建及从文库中分离筛选重要基因等研究工作的需要。
2.2 油茶种子表达基因分类在cDNA文库中,随机挑取2 327个克隆进行3′端单向序列测定。并对数据完整无N、X且序列长于200 bp的1 979条序列进行比对,筛选出了763条266种可确认的基因序列(谭晓风等,2005; 胡芳名等,2005),其中219条序列与NCBI的NR库和dbEST库中的其他物种cDNA同源性很高(分值≥200,相似性≥78%,同源序列长150 bp以上),225条序列同源性较高(分值≥100,相似性≥78%,同源序列长100 bp以上),319条序列的分值介于50~100之间,还有1 216条的同源性较低或为未知名基因序列。对2 327个克隆的序列分析结果见表 1。
|
|
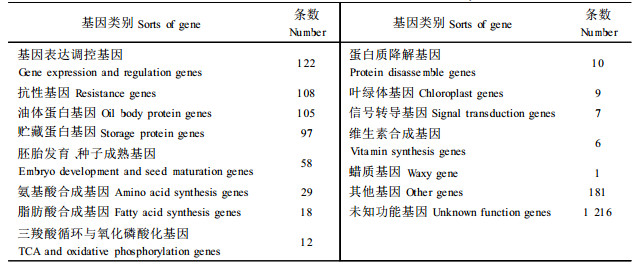
不同类型的基因其表达丰度不同,其中贮藏蛋白相关基因、基因表达相关基因、抗性相关基因、与发育相关的基因在种子中有高丰度表达。脂肪酸合成相关的基因属中丰度表达。有的基因只测到一个序列,属低丰度表达。油茶油脂转化高峰期与脂肪酸合成有关的代谢酶基因有18条序列,贮藏蛋白、油体蛋白基因有202条序列,抗病、抗逆性基因有108条序列等,未知功能的基因序列达1 216条。
2.3 与脂肪酸合成相关的基因在油脂转化高峰期,检测到与脂肪酸合成代谢相关的酶基因共有9种,分别为硬脂酰ACP脱饱和酶(SAD)基因、油酸脱饱和酶FAD2基因、FAD8基因、脂酰ACP硫酯酶(FatB1)基因、ACP基因、丙酰CoA合成酶基因、烯酰CoA水化酶基因、乙酰CoA转乙酰基酶基因、长链脂肪酸浓缩酶基因等。
长链饱和脂肪酸是由乙酰CoA在脂肪酸合成酶(FAS)系作用下合成的(卢善发, 2000)。该酶系以含泛酰巯基乙胺残基的酰基载体蛋白(acyl carrier protein, ACP)作为脂酰基载体。植物体内存在多个ACP同工酶(Heath et al., 1995), 如大麦(Hordeum vulgare)有3个, 分别为ACP-Ⅰ、ACP-Ⅱ和ACP-Ⅲ。ACP含有两类SH基团,分别由泛酰基巯基乙胺及半胱氨酰(Cys)残基提供。软脂酰ACP水解后生成软脂酸(16:0),软脂酸是长链饱和脂肪酸及不饱和脂肪酸的前体,在叶绿体中合成后运到胞质中,在内质网和线粒体内的脂肪酸延长体系作用下,分别与丙二酰CoA和乙酰CoA的残基连接而使碳链延长,生成硬脂酸(18:0)或更长碳链的饱和脂肪酸。在油茶种子cDNA文库中测到相关基因序列1条(rzots0_007062.y1.scf),为低丰度表达。与木麻黄(Casuarina glauca)的ACP序列比对分值为170,期望值2e-39, 相似性系数87%(152/174),具有很高的同源性; 可以认为所测的基因序列为油茶ACP基因序列。
硬脂酰-ACP脱饱和酶(stearoy1-acp desaturase, SAD)的主要作用是将硬脂酸脱氢后形成油酸(18:1),是不饱和脂肪酸形成的关键酶之一(Yukawa et al., 1996)。油茶cDNA文库测序获得了3条硬脂酰ACP脱饱和酶基因相关序列(rzots0_000185.y1.scf、rzots0_000468.y1.scf、rzots0_004461.y1.scf),属中丰度表达。以rzots0_004461.y1.scf序列比对分析的结果,与蓖麻(Ricinus communis)的SAD基因序列最为接近,分值383,期望值e-103,相似性84% (418/493)。该序列经2次测序,确定为油茶种子的SAD基因序列。
油酸脱饱和酶(FAD2)是植物产生多不饱和脂肪酸的关键酶,FAD2是专一的加氧酶(oxygenase)系统,它存在于内质网膜上,以1-酰基-2-油酰基-sn-甘油-3-磷酸胆碱为底物,在需氧条件下,以NADP+为辅酶生成多不饱和脂肪酸-亚油酸(cis 9, 12-二烯酸)和亚麻酸(cis 9, 12, 15-三烯酸),该基因已经在拟南芥(Arabidopsis thaliana)、落花生(Arachis hypogaea)、油菜(Brassics napus)等植物中克隆出来(Somerville et al., 1996; Wada et al., 1992; Tasaka et al., 1996)。油茶cDNA序列测定中获得3条油酸脱饱和酶相关基因序列,与FAD2ω-6脂肪酸脱饱和酶基因相关序列(rzots0_001934.y1.scf)1条、FAD8ω-3脱饱和酶基因相关序列(rzots0_001749.y1.scf)1条、FAD2 δ-12脱饱和酶基因相关序列(rzots0_007342.y1.scf)1条。其中FAD2ω-6脂肪酸脱饱和酶基因与芝麻(Sesamum indicum)FAD2的序列同源性很高,分值为176,期望值为4e-41, 序列相似性85% (188/221)。该序列经2次测序确认为油茶FAD2基因序列。
脂酰ACP硫酯酶(FatB1)是催化脂肪酸合成的关键酶之一,主要作用是终止脂肪酸链的延长(Heath et al., 1995)。脂酰ACP硫酯酶基因序列的获得对研究油茶脂肪酸合成FAS循环的终止,会起到积极的作用。所测油茶cDNA序列中共获得3条与FatB1基因相关序列(rzots0_000150.y1.scf,rzots0_000409.y1.scf,rzots0_007056. y1.scf),属中丰度表达。BLAST比对结果表明,与拟南芥等植物的FatB1基因的比对最好分值为128,期望值e-26,同源性81%。所测序列是否为脂酰ACP硫酯酶基因序列需进一步测定。
乙酰CoA转乙酰基酶(acetyl CoA acetyltransferase)催化2分子乙酰CoA转化成乙酰乙酰CoA,是一种硫解酶,参与类异戊二烯活体的合成(Daniel et al., 1991),是类固醇合成的启动步骤。在油茶cDNA中获得2条序列(rzots0_004412.y1.scf,rzots0_006155.y1.scf),对后1条序列进行BLAST比对,与巴西橡胶(Hevea brasiliensis)的乙酰CoA转乙酰基酶基因的相似性88% (408/461),分值494,期望值e-136,相似性很高,可确认所测序列为油茶种子乙酰CoA转乙酰基酶基因序列。
长链脂肪酸缩合酶(very-long-chain fatty acid condensing enzyme)是涉及长链脂肪酸生物合成的微粒体脂肪酸延长酶中4种活性酶的第一种,是决定长链脂肪酸的酰基链长度的延长酶中的主要活性酶(Millar et al., 1997)。测得油茶cDNA序列1条(rzots0_002724.y1.scf),属低丰度表达。BLAST比对结果,与拟南芥长链脂肪酸缩合酶基因的相似性81% (219/270),分值123,期望值5e-25,可认为所测序列为长链脂肪酸缩合酶基因序列。长链脂肪酸缩合酶具有种质特异性,其表达和特异性控制长链脂肪酸的生物合成。该酶基因的获得对研究油茶长链脂肪酸的合成,将会起到重要的作用。
2.4 蜡质基因蜡质基因是编码直链淀粉合成酶的关键基因,与种子中淀粉含量及直链淀粉与支链淀粉的比例有很大关系(耿俊丽等,2005), 也与长链脂肪酸和高级醇的形成有关。在油茶种子cDNA文库中测序得到1条蜡质基因序列(rzots0_000450.y1. scf),属低丰度表达。经与GenBank中核酸库比对,与金缕梅(Hamamelis vernalis)等植物的蜡质基因比对分值为109,期望值3e-21,同源性89%(85/95)。由于比对分值较低,所得序列需做进一步研究,以确认是否为蜡质基因。
2.5 油体蛋白基因植物种子中的油脂主要贮藏在亚细胞微粒形式的油体中。油体内部为液态的三酰甘油酯,外面覆盖着单层的磷脂分子和镶嵌蛋白质构成的半单位膜。膜中镶嵌的蛋白为油体相关蛋白,有oleosin、caleosin、steroleosin等(仇键等,2005)。油茶种子cDNA文库中测得油茶油质蛋白(oleosin)基因序列(rzots0_005927.y1.scf,rzots0_003552. y1.scf等)84条,脂体膜蛋白(lipid body membrane protein)基因(rzots0_004905. y1.scf等)17条,油体钙蛋白(caleosin) 3条(rzots0_001281.y1.scf等)及油体甾醇蛋白(steroleosin-B)1条等。其中油质蛋白与芝麻油质蛋白相似性最高,比对分值135,期望值e-28,相似性系数84%(152/180);脂体膜蛋白与胡萝卜(Daucus carta var. sativa)的比对分值为121,期望值2e-24,相似性系数87%(109/125);油体钙蛋白与芝麻的比对分值为58.0,期望值2e-05,相似性系数79%(119/149);油体甾醇蛋白与芝麻的比对分值139,期望值e-30,相似性系数80%(271/338)。以上高分值序列已进行2次测序,并确认所得序列为相应的基因序列。
2.6 贮藏蛋白基因不同类型的植物种子,贮藏物的种类和数量有所不同,贮藏物主要作为供应种子发芽及幼苗生长时所需养分及能量来源。测序获得油茶贮藏蛋白(β-globulin B)基因相关序列(rzots0_000424.y1.scf等)44条; 豆球蛋白(legumin)基因相关序列(rzots0_006926.y1.scf等)36条; 清蛋白(albumin)15条(rzots0_000145.y1.scf等); 谷蛋白(glutelin)2条(rzots0_001334.y1.scf,rzots0_002453.y1.scf)。BLAST的结果表明,油茶种子的贮藏蛋白基因与棉花的beta-globulin B基因的匹配分值都在50分以上,期望值< e-4,相似性系数86% (53/61)。油茶种子中的豆球蛋白基因与紫苏(Perilla frutescens)、棉花、欧洲栎(Quercus robur)等植物的legumin B基因的匹配分值都在60分以上,期望值< e-8,相似性系数89% (53/59)。油茶种子中清蛋白的基因序列与Bertholletia excelsia的比对分值为52.0,期望值0.002,相似性100%(26/26)。比对结果,总体分值较低,但局部相似性高,这与贮藏蛋白是小分子蛋白有关,贮藏蛋白的两端为高变区而中心为较小的保守区。
普通油茶种子油脂含量为20%~28%,种仁油脂含量40%~59%,富含不饱和脂肪酸和一定数量的蛋白质,为营养良好的蛋白质及油脂来源,油茶树被称为东方油橄榄,茶油可和橄榄油相媲美(Tan et al., 2004),其中贮藏蛋白基因中豆球蛋白是其主要蛋白之一(Tan et al., 2005)。在植物种子中贮藏蛋白质及油脂的细胞器分别为蛋白质体(protein body)和油体(oil body)。油茶种子EST中贮藏蛋白质基因、油脂贮藏基因的表达都是极高丰度的,同时检测到与脂肪运输相关的脂质转移蛋白基因(rzots0_003318.y1.scf)。这一结果与油茶种子正处油脂转化高峰期的生理特征是相吻合的。对这些基因进行研究,将对油茶种子脂类物质的产生、储存等过程有更深入的诠解。
2.7 基因表达调控相关的基因核糖体是蛋白质合成的场所,真核生物的核糖体由60S的大亚基和40S的小亚基组成。40S的小亚基由18S的rRNA分子和30种不同的核糖体蛋白质组成,60S大亚基由各一分子的5S rRNA, 5.8S rRNA, 28S rRNA和约45种蛋白质组成。核糖体常常分布在细胞蛋白质合成旺盛的区域,其数量和蛋白质合成强度有关。测序获得了26条核糖体结构基因如60S大亚基基因序列(rzots0_005968. y1.scf等)、40S小亚基基因序列(rzots0_001025.y1.scf等),属高丰度表达。经比对,大亚基基因序列与黄瓜(Cucumis sativus)的比对分值为325,期望值为6.00e-86,相似性系数83% (417/500);小亚基基因序列与鹰嘴豆(Cicer arietinum)的比对分值达309,期望值为3.00e-81,相似性系数85% (315/368)。可确定所得序列为相应的基因序列。
转录因子是基因表达必需的元件,对基因的表达起调控作用。所测序列中有MADS盒的转录因子基因(rzots0_007178.y1.scf),AP2包含域转录因子基因(rzots0_003331.y1.scf等),DRE捆绑转录因子基因(rzots0_000143.y1.scf),AG-基序捆绑蛋白基因(rzots0_004235.y1.scf)、F-box家族蛋白基因(rzots0_000916.y1.scf)及翻译起始因子基因(rzots0_001941.y1.scf等),这些因子对基因的表达起重要作用。
2.8 发育有关基因油茶种子的生长、发育与成熟过程受多种基因的调节控制。在油茶种子cDNA文库中获得16种58条与油茶种子生长、胚胎发育及成熟相关的基因序列,这些序列与核酸数据库中其他物种相关序列有很高或较高的同源性。其中脱落酸应答蛋白(abscisic acid response protein)基因序列22条, 成熟控制蛋白(ripening regulated protein)和翻译控制肿瘤蛋白(translationally controlled tumor protein,TCTP)基因序列各6条, 胚胎特化蛋白(embryo-specific protein)和乙烯应答因子包含域蛋白(AP2/ERF-domain protein)基因序列各4条,为高丰度表达; 乙烯应答因子(ethylene response factor)基因序列3条,种子成熟蛋白(seed maturation protein)、细胞分化蛋白(cell differentiation protein)及脱水素(dehydrin)基因序列各2条,属中等丰度表达; 而成熟相关蛋白(ripening-related protein)、胚珠发育蛋白(ovule development protein)、生长控制因子(growth-regulating factor)、休眠相关蛋白(dormancy related protein)、衰老相关蛋白(senescence-associated putative protein)和脱水应答蛋白等基因序列各测到1条,属低丰度表达(谭晓风等,2005)。
以上所测cDNA序列分析,基本反映出油茶种子与胚胎发育和种子成熟相关基因的阶段性表达情况,油茶种子的发育已进入到种子成熟和衰老状态,与成熟和衰老关系极为密切的植物激素脱落酸、乙烯进入快速形成期,脱水素基因及与衰老和休眠相关的基因也已表达。相关分析另文发表。
2.9 抗逆相关基因植物体内存在着抵御外界不良环境、外源病虫害等生物侵袭的抗性基因,它能加强和改善植物自身的防御系统,有效地抵御病虫害的侵袭,增强对逆境的适应能力。在油茶种子cDNA文库中获得23种108条与油茶种子抗性相关的基因序列,其中核糖体灭活蛋白(ribosome-inactivating protein)基因35条、类金属硫蛋白(metallothionein-like protein)基因18条、过氧化物酶(peroxidase)基因11条、热激蛋白(heat shock protein)基因9条、查尔酮合酶(chalcone synthase)基因5条,属高丰度表达; 铜锌超氧化物歧化酶(copper/zinc-superoxide dismutase)基因、植物外源凝集素(lectins)基因及富含甘氨酸蛋白(glycine rich protein)基因序列各3条,过氧化氢酶(catalase)基因、防御蛋白(defensin protein 1)基因、抗性基因簇(resistance gene cluster)、木质素蛋白(xylogen protein)基因、环氧化物水解酶(epoxide hydrolase)基因序列各2条,属中等丰度表达; 另有8种抗性基因二氢黄酮异构酶(rapid cycling dihydroflavonone isomerase)基因、非细胞自主的同源热激蛋白(non-cell autonomous heat shock cognate protein virus)、抗病毒基因位点(resistance gene locus)、类黄酮转葡萄糖基酶(isoflavonoid glucosyltransferase)基因、黄酮醇合酶(flavonol synthase)基因、耐盐蛋白基因(salt tolerance protein 1)、伸展蛋白基因(extensin)及苯丙氨酸氨基裂解酶(phenylalanine ammonia-lyase)基因各1条序列,属低丰度表达。油茶种子中大量抗性基因的表达与其生理过程及受到外源生物的侵袭有密切的关系。
2.10 其他表达基因油茶种子cDNA文库中还有氨基酸合成(amino acid synthesis)基因序列29条,重金属解毒蛋白基因序列18条,三羧酸循环与氧化磷酸化(TCA and oxidative phosphorylation)基因序列12条,蛋白质降解(protein disassemble)基因10条,叶绿体(chloroplast)基因序列9条,信号转导(signal transduction)基因序列7条,维生素合成(vitamin synthesis)基因序列6条,及参与肽链折叠的蛋白因子基因、肌动蛋白基因、ACCase基因、RCMIT基因、TITP基因、ARF、UBC基因、钙调素基因等及许多功能未知的基因序列,在此不一一详述。
3 结论与讨论不同植物种子合成的油脂种类和数量差异很大。油茶种子能合成大量有利于人类健康的不饱和脂肪酸(雷治国等, 2003)。为开发油茶的优质基因资源,克隆相关基因并进行遗传改良,作者利用油茶近成熟种子为材料建立cDNA文库,随机对其中的2 327个克隆进行测序,建立了油茶近成熟种子的EST文库。
研究发现油茶近成熟的种子中脂肪酸合成代谢较强,测序获得了SAD基因、FAD2基因、FAD8基因、ACP基因、FatB1基因等9种18条序列。基本上反映出近成熟油茶种子脂肪酸合成代谢的主要基因表达情况。但乙酰CoA羧化酶等关键酶未找到相应的基因序列,需做进一步研究。
油体蛋白基因、豆球蛋白基因、贮藏蛋白基因的mRNA序列数约占总mRNA序列数的10.5%,是主要的表达基因。反映油茶种子正处油脂转化高峰期。在油茶种子中发现有农作物水稻(Oryza sativa)等种子胚中的谷蛋白基因序列,但没有发现醇溶蛋白基因序列。油茶种子中存在多种核糖体蛋白基因、核糖体RNA基因及其他与蛋白翻译有关的基因,说明在油茶种子的油脂转化过程中同时有大量蛋白质的合成,而蛋白质降解基因的表达,可能与部分蛋白质转化成油脂有关。呼吸作用相关基因的表达与油茶种子正处油脂转化高峰期相适应,通过呼吸代谢为油脂合成提供原料。与发育和成熟相关的基因大量表达,说明油茶种子发育到这一阶段,正在为种子成熟进入休眠做准备。
油茶种子中核糖体灭活蛋白基因、金属硫蛋白基因、过氧化物酶基因、热激蛋白基因、查尔酮合酶基因、抗病毒基因、过氧化物酶基因、铜锌超氧化物歧化酶基因、外源凝集素基因、防御蛋白基因、干旱应答蛋白基因、胁迫相关蛋白基因、耐盐蛋白及热激蛋白基因等耐环境胁迫基因均有表达。这与油茶种子具有较强的抗病和耐环境胁迫的能力相适应。
油茶近成熟种子中,还有一些新基因特异性表达,但大多数呈中低丰度状态,如噻唑生物合成酶基因。多数只测到1条序列,如Ras-相关蛋白基因、Profilin基因、Synptobrevin相关蛋白基因、Fibrillarin基因、Remorin基因等。上述基因序列的获得,为以后油茶油脂合成代谢过程及各种关键酶基因全序列的进一步研究,奠定了物质和技术基础。
陈永忠, 张智俊, 谭晓风. 2005. 油茶优良无性系的RAPD分子鉴别. 中南林学院学报, 25(4): 44-49. |
黄永芳, 陈锡沐, 雷治国, 等. 2004. 油茶种质资源RAPD分析Ⅰ:DNA提取和PCR扩增条件建立. 河南农业科学, (12): 23-26. |
黄永芳, 吴雪辉, 陈锡沐, 等. 2005. 引物对油菜种质资源聚类分析结果的影响. 河南农业科学, (10): 39-43. |
何方, 胡芳名. 2004. 经济林栽培学. 2版. 北京: 中国林业出版社, 278-279.
|
胡芳名, 谭晓风, 石明旺, 等. 2004. 油茶种子cDNA文库构建. 中南林学院学报, 24(5): 1-4. DOI:10.3969/j.issn.1673-923X.2004.05.001 |
胡芳名, 谭晓风, 仇键, 等. 2005. 油茶种子中主要表达的储藏蛋白相关基因. 中南林学院学报, 25(4): 24-26. DOI:10.3969/j.issn.1673-923X.2005.04.005 |
耿俊丽, 胡芳名, 张党权. 2005. 蜡质基因研究进展. 中南林学院学报, 24(5): 101-104. |
江昌俊. 1995. 欧洲油菜花粉cDNA文库的构建. 安徽农业科学, 23(3): 4-8. |
雷治国, 黄永芳, 何会蓉. 2003. 油茶及其种质资源研究进展. 经济林研究, 21(4): 123-125. DOI:10.3969/j.issn.1003-8981.2003.04.041 |
刘长征, 张正舫, 张国珍, 等. 1995. 玉米叶片cDNA文库构建及Cah基因克隆. 山东农业大学学报, 26(4): 447-452. |
骆蒙, 贾继增. 2001. 植物基因组表达序列标签(EST)计划研究进展. 生物化学与生物物理进展, 28(4): 494-497. DOI:10.3321/j.issn:1000-3282.2001.04.014 |
卢善发. 2000. 植物脂肪酸的生物合成与基因工程. 植物学通报, 17(6): 481-491. DOI:10.3969/j.issn.1674-3466.2000.06.001 |
裴凌鹏, 李伟, 黄勤妮. 2003. EST在植物代谢研究中的应用. 北京联合大学学报:自然科学版, 17(3): 83-86. DOI:10.3969/j.issn.1005-0310.2003.03.020 |
仇键, 谭晓风. 2005. 植物种子油体及相关蛋白研究综述. 中南林学院学报, 24(5): 96-100. |
萨姆布鲁克J, 拉塞尔D W. 1999.分子克隆实验指南.黄培堂, 等译.2版.北京: 科学出版社, 2-449
|
谭晓风, 胡芳名, 谢禄山, 等. 2005. 油茶近成熟种子表达的发育相关基因及其分析. 中南林学院学报, 25(4): 17-23. DOI:10.3969/j.issn.1673-923X.2005.04.004 |
张光祥, 马正强. 2003. 目的全长cDNA合成及克隆技术进展. 生物技术通报, 14(5): 1-5. DOI:10.3969/j.issn.1002-5464.2003.05.001 |
张云, 黄儒珠, 刘大林, 等. 2003. 油茶随机扩增多态DNA条件的研究. 福建林业科技, 30(2): 8-11. |
钟华, 谢婷婷, 刘中来. 2003. EST的应用现状与前景. 生物技术, 13(1): 32-33. DOI:10.3969/j.issn.1004-311X.2003.01.023 |
中国科学院植物研究所. 1971. 中国高等植物图鉴:2册. 北京: 科学出版社, 856.
|
Chang S, Puryear J, Cairney J. 1993. A simple and efficient method for isolating RNA from pine trees. Plant Mol Biol Repotr, 11: 113-116. DOI:10.1007/BF02670468 |
Daniel J Petersen, George N Bennett. 1991. Cloning of the Clostridium acetobutylicum ATCC 824 acetyl coenzyme A acetyltransferase (Thiolase; EC 2.3.1.9) gene. Applied and Environmental Microbiology, 57(9): 2735-2741. |
Heath R J, Rock C O. 1995. Enoyl-acyl carrier protein reductase (fabI) plays a determinant role in completing cycles of fatty acid elongation in Escherichia coli. J Biol Chem, 270: 26538-26542. DOI:10.1074/jbc.270.44.26538 |
Millar A A, Kunst L. 1997. Very-long-chain fatty acid biosynthesis is controlled through the expression and specificity of the condensing enzyme. Plant J, 12(1): 121-131. DOI:10.1046/j.1365-313X.1997.12010121.x |
Somerville C, Browse J. 1996. Dissecting desaturation. Trendsin Cell Biol, 6: 148-153. DOI:10.1016/0962-8924(96)10002-7 |
Tan Xiaofeng, Hu Fangming, Shi Mingwang, et al. 2004. The main expressed genes during the transformation peak of oil in seeds of Camellia oleifera. The Second International Forum on Post-Genome Technologies, Nanjing, China: 98. |
Tan Xiaofeng, Zhang Dangquan, Qiu Jian, et al. 2005. Separation and bioinformatic analysis of oleosin genes in seeds of Camellia oleifera. The Third International Forum on Post-Genome Technologies, Guilin, China: 90-91. |
Tasaka Y, Gombos Z, Nishiyaa Y, et al. 1996. Targeted mutagenesis of acyL-lipid desaturase in synichocystis: evidence for the important roles of polyunsaturated membrane lipids in growth respiration and photosynthesis. EMBOJ, 15(23): 6416-6425. DOI:10.1002/j.1460-2075.1996.tb01033.x |
Wada H, Gombos Z, Sakamoto T, et al. 1992. Genetic manipulation of the extent of desaturation of fatty acid in membrane lipids in the Cyanobacterium synecocyst is PCC6803. Plant Cell Physiol, 33(5): 535-540. |
Yukawa Y, Takaiwa F, Shojik K, et al. 1996. Structure and expression of two seed-specific cDNA encoding stearoyl-acylcarrier protein desaturase from sesame, Sesamum indicum. Plant Cell Physi, 37(2): 201-205. |