 2006, Vol. 42
2006, Vol. 42文章信息
- 王娟, 马钦彦, 杜凡.
- Wang Juan, Ma Qinyan, Du Fan.
- 云南大围山国家级自然保护区种子植物区系多样性特征
- Flora Diversity Characteristics of Seed Plants of Dawei Mountain National Nature Reserve in Yunnan Province, China
- 林业科学, 2006, 42(1): 7-15.
- Scientia Silvae Sinicae, 2006, 42(1): 7-15.
-
文章历史
- 收稿日期:2004-06-24
-
作者相关文章


2. 北京林业大学资源与环境学院 北京 100083
2. College of Resources and Environment, Beijing Forestry University Beijing 100083
大围山国家级自然保护区位于北回归线以南,东经103°20′—104°03′,北纬22°35′—23°07′,面积153.7 km2,四周高山阻挡,仅东南面靠狭窄的红河谷口(海拔76.4 m)通过越南到北部湾。这种特殊的槽式地形极易接纳和滞留北部湾东南季风暖湿气流。该地区雨热丰富,夏天高温多雨,冬季暖湿多雾,年降雨量1 700~1 900 mm,每年的1—5月云南春旱时,该区降雨量仍然达到420 mm以上;年均温22.6 ℃,最冷月(1月)均温15.2 ℃,最热月(7月)均温27.7 ℃,极端低温多年平均值为5.8 ℃,大于10 ℃的年积温8 246.2 ℃,全年无霜。区内最低处海拔225 m;最高峰大尖山海拔2 365 m,相对高差达2 140 m,随海拔升高依次分布湿润雨林、山地雨林、湿性季风常绿阔叶林、山地苔藓常绿阔叶林和山顶苔藓矮林等植被类型。
植物区系的多元性作为生物多样性表征之一,其组成、发生、发展、迁移、演化从多方面反映该区域植物多样性的特性(蒋有绪等,2002)。大围山位于滇东南云南高原向南倾斜面南缘的一列断块山地上,处于大陆东南亚热带北缘热带生物区系向云南高原亚热带生物区系过渡的地带,同时又处于‘田中线’(李锡文等,1992)南端附近东亚区系两大成份相汇处。该区在植被地理、植物区系地理和生物多样性保护研究上具有较高的科学价值和实践意义。19世纪以来,中外学者到该区进行植物标本采集和考察,其成果主要集中反映在《云南植物志》、《云南种子植物名录》等专著中。1997年以来,西南林学院对大围山自然保护区进行了全面的综合考察。本文系统研究了该区种子植物区系多样性特征,可为深入探讨滇东南与滇南、滇西南植被地理和植物区系地理内在联系及差异提供依据。
1 研究方法依据野外标本采集、样地调查和相关文献资料(吴征镒, 1991;税玉民, 2003)建立大围山种子植物区系数据库。采用植物区系地理的理论和方法(王荷生,1992;吴征镒,1991;吴征镒等,2003;李锡文,1996),分析大围山种子植物区系成分及其多样性特征。
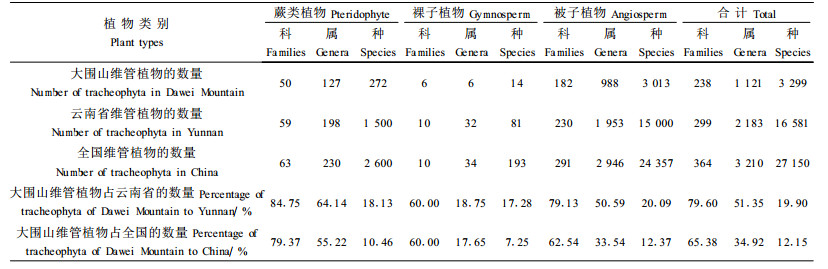
2 结果与分析 2.1 植物资源多样性大围山国家级自然保护区有野生种子植物188科994属3 027种及变种,种属比3.05。其中裸子植物6科6属14种,双子叶植物160科787属2 491种,单子叶植物22科201属522种。平均种密度高达19.69种·km-2,高于滇西南铜壁关自然保护区的11.32种·km-2(西南林学院等,2004)和滇南西双版纳自然保护区的1.4种·km-2(李延辉等,1996)。
大围山保护区面积仅为云南总面积39.4万km2的1/2 000,却汇集了云南省蕨类植物的18.13%,裸子植物的17.28%,被子植物的20.09%(郭辉军等,1998)(表 1)。维管植物的科、属、种数分别占全国总科、属、种的65.38%、34.92%、12.15%。这充分说明大围山植物物种多样性极为丰富,物种分布密度高,在生物多样性保护中占有重要地位。
|
|
该区188个科的种子植物中,含90种以上的科有5个,其中种数最多是茜草科(Rubiaceae)(47:74) (属数:种数, 以下同),以下依次为兰科(Orchidaceae) (61:159)、菊科(Compositae) (53:123)、禾本科(Gramineae) (65:121)、樟科(Lauraceae) (13:95)。它们仅占该区总科数的2.7%,所含属数和种数却占总属数的24.0%和总种数的22.2%。兰科、菊科、禾本科是世界性分布的几个含数千种以上的大科,在该区也有极为丰富的属种。茜草科和樟科是以热带分布为主的大科,前者热带性质较强,世界约含6 000种,中国约有550种,本区有全国种数的31.6%;后者是以热带、亚热带地区为主分布的科,世界约含2 000种,中国种数约430种,本区有全国种数的22.1%。含30~89种的有壳斗科(Fagaceae)(5:77)、山茶科(Theaceae)(9:62)、桑科(Moraceae)(6:53)、苦苣苔科(Gesneriaceae) (17:50)、紫金牛科Myrsinaceae (5:46)、野牡丹科(Melastomataceae) (16:44)、番荔枝科(Annonaceae) (11:44)等25个科,这些科中除蔷薇科(Rosaceae)(22:81)、莎草科(Cyperaceae) (10:46)、唇形科(Labiatae) (21:45)、玄参科(Scrophulariaceae) (19:37)为世界广布科,百合科(Liliaceae)为温带性质的科外,其余20个科均是热带性质的科。
以上两类科占本区总科数的16.0%,它们的属和种分别占总属、种数的56.2%、62.6%,是该植物区系的重要成分和数量上的优势科,其热带性质的科占83.33%,反映该植物区系强烈的热带性质。
样方调查结果显示:重要值大于1的壳斗科、樟科、木兰科(Magnoliaceae)、魏香科(Styracaceae)、山茶科、金缕梅科(Hamamelidaceae)、桦木科(Betulaceae)、杜英科(Elaeocarpaceae)、五加科(Araliaceae)、大戟科(Euphorbiaceae)、蔷薇科(Rosaceae)、石楠科(Ericaceae)、无患子科(Sapindaceae)、山榄科(Sapotaceae)、龙脑香科(Dipterocarpaceae)、紫树科(Nyssaceae)、桑科、紫金牛科的17个科中,除世界分布的蔷薇科和温带性质的桦木科、石楠科、珙桐科(Nyssaceae)外,其余的均为热带性质的科,也反映了该植物区系的热带性质。以不同温度性质的科进行统计,热带性质的科是温带性质科的两倍多,图 1。

|
图 1 大围山国家级自然保护区种子植物科级分布区类型比例图(数字2~14代表的分布区类型见表 3) Fig. 1 The percentage of seed plants family distribution areal-types in Dawei Mountain National Nature Reserve (The numbers from 2 to 14 of distribution areal0types represent refer to Tab. 3) C-世界分布科Cosmopolitan;Tr-热带性质的科From 2 to 7;Tm-温带性质的科from 8 to 14;Ch-中国特有分布Endemic to China. |
以上分析说明大围山种子植物区系以热带性质的科占绝对优势,在热带性质的科中又以主产热带、分布区扩展到亚热带甚至温带的科占有较大的比例。温带性质的科也占有一定比例反映了该植物区系成分的热带北缘性质和区系组成的边缘性。这与其位于热带亚洲边缘,且处于热带东南亚到云南亚热带高原过渡位置相吻合。
2.2.2 属级区系成分多样性特征1) 属组成多样性特征根据所含种数将994个属分为较大属(含21种以上),多种属(10~20种),中等属(含5~9种),寡种属(含2~4种),单种属(表 2)。含10种以上的较大属和多种属计58个,占总属数的5.84%。其中世界广布7属,温带性质7属,热带性质达44属,也反映了植物区系的热带性质。群落调查结果也显示,上层乔木层多由杜英属(Elaeocarpus)、冬青属(Ilex)、石栎属(Cinnamomum)、樟属(Lithocarpus)、栲属(Castanopsis)、琼楠属(Beilschmiedia)等属的种类组成;中下层乔木以榕属(Ficus)、木姜子属(Litsea)、山矾属(Symplocos)、茶属(Camellia)、胡椒属(Piper)、柃属(Eurya)等属为主要组成成分;灌木以粗叶木属(Lasianthus)、紫金牛属(Ardisia)、乌饭树属(Vaccinium)、悬钩子属(Rubus)等种类较多;草本则以秋海棠属(Begonia)、楼梯草属(Elatostema)等种类较为丰富,此外还有大量的附生植物,如瓜馥木属(Fissitigma)、崖藤属(Tetrastigma)较多。单种属和寡种属极为丰富,两者共占总属数83.7%,所含种数占46.15%。它们是构成大围山植物区系多元化的主要原因之一,表明大围山处于热带区系的北缘和亚热带区系南缘的某些扩散式分布特征。大围山的种属系数(种/属)3.05,远低于横断山脉(6.0)、青藏高原高寒区(5.27)、滇黔桂(4.28)等地区(李锡文,1995),因此具有属内种系相对贫乏,分化程度较弱的特点。
|
|
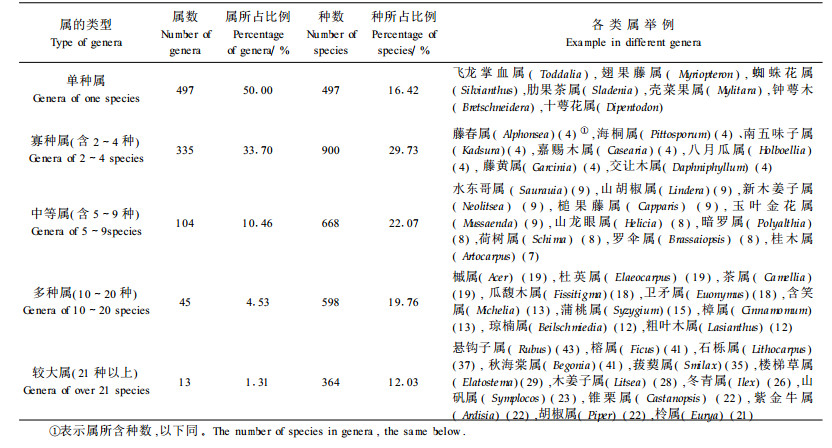
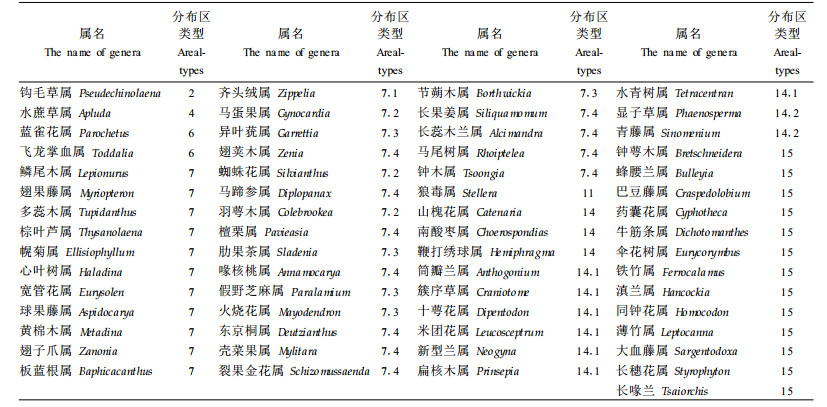
2) 属分布区类型多样性特征中国种子植物属15个分布区类型31个变型中(吴征镒,1991),大围山有15个类型19个变型(表 3),反映其种子植物区系联系的广泛性和复杂性。①世界分布占总属数的3.92%,含230种,占总种数的7.60%。以草本和灌木为主,如早熟禾属(Poa)、半边莲属(Lobelia)、茄属(Solanum)、沟酸浆属(Mimulus)、酸横属(Rumex)等的种类。其中很多几乎只见于路边、荒坡、草丛,在森林群落中并不多见。②热带分布属及其多样性特征热带分布的6种类型在大围山都存在,共计698属,占该区非世界属数的73.1%;含2 150种,占总种数的76.9%,其中热带分布属数与温带分布属数的比值为Tr/Tw=3.05。反映了大围山种子植物区系强烈的热带性质及热带植物的丰富性。其中热带亚洲分布及其变型最多,计289属,占该区非世界属数的30.3%;含612种,占总种数的21.89%。这一带本身就是热带亚洲的北缘部分,显示该植物区系与热带亚洲植物区系的亲缘关系(姜汉侨,1980;吴征镒等,1987),特别是中南半岛区系的紧密联系。大围山分布有最典型的热带亚洲属,如龙脑香属(Dipterocarpus) (1)、婆罗双属(Shorea) (1)、隐翼属(Crypteronia) (1)、八宝树属(Duabanga) (1)、无忧花属(Saraca) (1)、白颜树属(Gironniera) (1)、子楝树属(Decaspermum) (1)、假山龙眼属(Heliciopsis) (1)、黄内楠属(Actinodaphne) (5)、木奶果属(Baccaurea) (1)、木桂属(Artocarpus) (7)、木莲属(Manglietia) (6)等,构成大围山热带森林群落的主要上层乔木成分。这一类型中单型属比例较高,计31属,占该类型的10.73%。
|
|

泛热带分布型及变型占该区非世界属数的17.70%,含788种,占总种数28.17%,虽然其属的数量仅次于热带亚洲分布属,但所含种数则远较热带亚洲分布属多,其中很多是森林群落的固有成分,如云实属(Caesalpinia) (2)、石豆兰属(Bulbophyllum) (14)、虾脊兰属(Calanthe)(12)、柞木属(Xylosma) (2)、罗汉松属(Podocarpus) (4)、卫矛属(Euonymus)(18)、粗叶木属(Lasianthus) (12)、山黄皮属(Randia) (2)、琼楠属(Beilschmiedia) (12)、鹅掌柴属(Schefflera) (14)、杜英属(Elaeocarpus) (19)、菝葜属(Smilax) (33)、白粉藤属(Cissus) (5)、山矾属(Symplocos) (23)、赤贞桐属(Clerodendrum) (16)、崖豆藤属(Millettia)(9)、冷水花属(Pilea) (14)、紫金牛属(Ardisia) (18)、冬青属(Ilex) (26)、李榄属(Linociera) (1)、榕属(Ficus) (35)、秋海棠属(Begonia) (39)等的种类占据着热带森林的各个层次。该类型单型属数量极少,仅禾本科的钩毛草属(Pseudechinolaena)1属。
热带亚洲和热带美洲间断分布占该区非世界属数的2.72%。它们常常出现在亚热带性质的森林群落中,如檬果樟属(Caryodaphnopsis) (2)、木姜子属(Litsea) (24)、楠属(Phoebe) (5)、柃木属(Eurya) (21)、水冬哥属(Saurauia) (9)、无患子属(Sapindus) (2)、泡花树属(Meliosma) (10)、山香圆属(Turpinia) (6)、槟榔青属(Spondias) (1)、山柳属(Clethra) (2)、萝芙木属(Rauvolfia) (1)、猴欢喜属(Sloanea) (4)等。
旧世界热带分布及其变型占该区非世界属数的9.21%,如暗罗属(Polyalthia) (7)、紫玉盘属(Uvaria) (3)、青藤属(Illigera) (7)、地不容属(Stephania) (5)、海桐属(Pittosporum) (4)、蒲桃属(Syzygium) (13)、谷木属(Memecylon) (2)、金锦香属(Osbeckia) (4)、竹节树属(Carallia) (2)、闭花木属(Cleistanthus) (1)、血桐属(Macaranga) (6)、野桐属(Mallotus) (10)、橄榄属(Canarium) (6)、八角枫属(Alangium) (6)、杜茎山属(Maesa) (14)、狗牙花属(Ervatamia) (3)等。
热带亚洲至热带大洋州分布及其变型占该区非世界属数的6.70%。较重要的属有假鹰爪属(Desmos) (2)、樟属(Cinnamomum) (12)、风吹楠属(Horsfieldia) (1)、山龙眼属(Helicia) (6)、五桠果属(Dillenia) (2)、桃金娘属(Rhodomyrtus) (1)、野牡丹属(Melastoma) (4)、猫乳属(Rhamnella) (1)、嘉榄属(Garuga) (1)、米仔兰属(Aglaia) (2)、红椿属(Toona) (2)、假山萝属(Harpullia) (1)、桄榔属(Arenga) (1)、开唇兰属(Anoectochilus) (5)、隔距兰属(Cleisostoma) (3)、石仙桃属(Pholidota) (5)等。它们多数是热带森林群落中的固有成分,表明该植物区系与热带大洋州植物区系的联系。
热带亚洲至热带非洲分布及其变型占该区非世界属数的6.49%;含132种,占总种数的4.72%。较为常见和重要的属有飞龙掌血属(Toddalia) (1)、浆果楝属(Cipadessa) (1)、朱果藤属(Roureopsis) (1)、常春藤属(Hedera) (1)、铁仔属(Myrsine) (3)、牛角瓜果(Calotropis) (1)、杆柳属(Periploca) (3)、水团花属(Adina) (1)、雪花属(Argostemma) (2)、鹰爪花属(Artabortys) (4)等。它们常见于大围山海拔较低的热带森林群落中,少量见于海拔较高的温湿森林群落,部分出现于路边、荒坡、草地等非林的次生林带。
综上所述,大围山植物区系中热带分布属占优势,即热带成分诸类对整个区系的贡献远大于温带植物分布,在热带分布的属中,又明显以热带亚洲分布居首位,占热带分布属的41.40%,且有典型的热带亚洲属的存在,故又带有明显的热带亚洲植物区系特征。
③ 温带分布属的多样性特征温带分布属占该区非世界属数的23.98%;含616个种,占总种数的22.02%。这一数字远远低于中国范围内同类属所占的比例(40.87%),但明显高于云南南部西双版纳国家级自然保护区同类属的比例(15.31%)。就全国而言,大围山纬度偏南,又深受东南季风和西南季风暖湿气流影响,气候类型总体上属北热带类型,所以植物区系中的温带成分较整个中国的温带成分大大降低。其次,与西双版纳相比,大围山因地理位置和地形地势差异,海拔相对高差大,又属中山峡谷地貌,导致水热气候、生境分化,植物类型更为复杂,大量出现云南其他南部地区不存在的温带成分。因此,大围山的温带成分比西双版纳高许多,充分表明该植物区系成分的多样性和复杂性。
温带性质的属中,数量最多的东亚分布型及其两个变型有84属,占该区非世界分布属的8.80%,占本区温带属的36.68%,其中以全东亚分布占优势达43属,占整个东亚分布及其变型的51.19%,中国-喜马拉雅分布亚型有31属,占整个东亚分布及其变型的36.91%。居第二位的是北温带分布及其变型共有76属,占该区非世界属数的7.96%和占本区温带属的33.19%。这种情况与整个中国种子植物属中,前者的比例是9.93%,后者是10.03%的情况刚好相反。这一细微的差异反映出该区与东亚,尤其与东喜马拉雅的关系更紧密的区域特点。
东亚与北美洲间断分布及其变型占该区非世界属数的3.87%。较常见的有木兰属(Magnolia) (1)、五味子属(Schisandra) (4)、栲属(Castanopsis) (22)、石栎属(Lithocarpus) (37)、山核桃属(Carya) (1)、灯台树属(Thelycrania) (1)、紫树属(Nyssa) (1)、葱木属(Aralia) (6)、人参属(Panax) (2)、南烛属(Lyonia) (4)、马醉木属(Pieris) (1)、岩扇属(Shortia) (1)、流苏树属(Chionanthus) (2)、木樨属(Osmanthus) (6)、鼠刺属(Itea) (5)、八角属(Illicium) (6)等。它们是季风常绿阔叶林的建群和共建成分,反映出该区森林植被与东亚和北美洲的密切联系。
中亚分布和地中海区、西亚至中亚分布及其变型,前者在大围山没有分布,后者仅有木犀榄属(Olea),含短柄木樨榄(Olea brevipes)、尖叶木樨榄(Olea ferruginea)、云南木樨榄(Olea yunnanensis)3种,均生长于林下生境中,已经不再是干旱环境的物种。反映了大围山植物区系与干旱环境联系不密切的事实。
2.3 与云南热带植物区系的比较“地理成分是按各分类单位(特别是种)的现代分布区来分析的。可以归为若干个分布区类型Areal_type,然而它并不反映植物区系的发生”1)。因此,对属的分布区类型所做的统计仅仅给出其现代分布的概念。进行区系分析的目的在于了解地理成分与生境等的吻合程度,与邻近地区的差异程度,以及在多大程度上反映该区系的实质,尽可能地提供关于发生成分的概念,最终达到明确该区系起源、发生和发展。因此,将该区系属的分布区类型组成与中国、云南热带(李锡文,1995)的区系组成作一比较分析(图 2),探讨该区生物多样性发生的可能的历史原因。
1) 吴征镒.1984.植物区系地理学教学大纲:上下册.云南省植物学会印,17页

大围山与云南热带种子植物属的分布区类型构成相比,除东亚分布型比例较高外,其余几个分布区类型所占比例基本一致,这表明在云南热带的东南部、南部、西南部3片区域中,大围山作为滇东南较为典型的代表,其东亚分布型所占比例(8.46%)较南部、西南部要高,如西双版纳4.73%(李延辉等,1996),南滚河自然保护区4.34%(西南林学院等,2004),证实了大围山植物区系与东亚植物区系的联系紧密。此外,大围山与整个中国种子植物属的分布区类型构成相比,其热带成分比例明显较高,而温带成分比例较低,这与其所处的地理位置是一致的。中国境内有中国特有属257属,区内的中国特有属占中国这一类型属数的10.89%,其比例相对较低,这主要是由于大围山位于中国的西南部,与越南接壤,许多中国大陆其他地区的中国特有属很少分布到这一地区所致。
上述分析显示大围山是多种区系成分的交汇地带,但其热带成分在区系组成中明显占居优势地位,反映了该区系的热带性质,同时又与东亚区系成分,特别是与东喜马拉雅的植物区系有着紧密联系。反映干旱生境的中亚分布和地中海区、西亚至中亚分布极少甚至不存在,故该植物区系与干旱生境联系不密切。
2.4 植物区系的古老性 2.4.1 科级区系成分的古老性大围山的科组成中,单型科有肋果茶科(Sladeniaceae)、伯乐树科(Bretschneideraceae)、马尾树科(Rhoipteleaceae)、水青树科(Tetracentraceae)、大血藤科(Sargentodoxaceae)、十萼花科(Dipentodontaceae),它们在分类地位上十分孤立或起源上较为古老。其中大血藤科为中国特有科;水青树科、十萼花科为东亚特有科;肋果茶科则为热带亚洲的特有科;伯乐树科为缅甸、泰国至中国西南特有的科;马尾树科为越南(或中南半岛)至华南的特有科。表明该植物区系的古老性和在地质历史上与上述地区的联系。此外,在大围山较为古老的科及其种属大量存在,如多心皮的木兰科、八角科、番荔枝科等和柔荑花序类的如金缕梅科、壳斗科、荨麻科(Urticaceae)、胡桃科(Juglandaceae)等在大围山均可找到充分的代表。木兰科是最古老的被子植物之一,有6属22种,且以较为原始的云南拟单性木兰(Parakmeria yunnannensis)和木莲属、含笑属(Michelia)等的种类占多数。金缕梅科也是一个古老的科,在该区有8属11种,较为原始的几个属为典型东亚分布的蜡瓣花属(Corylopsis)(1)、马蹄荷属(Exbucklandia) (2)、壳菜果属(Mytilaria) (1)、红花荷属(Rhodoleia) (2)等,它们都是森林植被的建群种或重要组成成分。番荔枝科(44种)、壳斗科(77种)、荨麻科(78种)等也有众多的种类是该区数量上的优势科。它们的存在均显示大围山植物区系具有古老性特征。
2.4.2 属级区系成分的古老性大围山植物区系组成中单种属和寡种属极为丰富,两者共占总属数83.7%。其中单型属61属(表 4)和少型属100属,占总属数的16.40%。在61个单型属中,热带分布有35属,占55.56%,而温带亚洲分布仅1属,东亚分布12属,中国特有分布13属。从大围山东亚成分属的组成来看,在84个东亚分布属中,仅单型属和少型属有33属,占该区内东亚分布属的39.29%, 如此高的比例显示了该区东亚区系成分的古老性。大围山极为丰富的单种属和寡种属、高比例的单型属和少型属、裸子植物和蕨类植物及其古老类型的存在都反映了该区域植物区系的多元化和古老性。
|
|
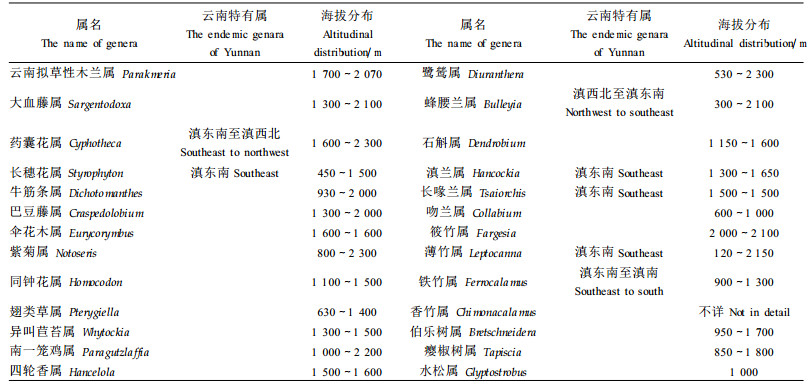
大围山植物物种多样性的另一特征是具有显著的特有性。区内26个中国特有属,分属21科,共31种,单型属14属和少型属6属共占76.9%。其中有7个属仅分布于云南(表 5)。高比例的单型属和少型属,其中一些属所代表的科在系统发育上相对原始或较为孤立,以及分布有中新世地层中的化石植物瘿椒树属(Tapiscia) (路安民,1999),均证实该区中国特有属的古老性。此外,生活型统计,这26个中国特有属中木本属占52%,草本属占48%,与中国特有属中草本属约占66%相比,其木本属比例较高,也反映大围山中国特有属植物的残遗性。大围山古老的特有属丰富,这与应俊生等(1994)认为滇东南-桂西为古特有属分布中心的论断是一致的。当然,上述中国特有属中有的可分布到邻国,如箭竹属分布到缅甸北部和东喜马拉雅地区,为横断山—喜马拉雅成分,东喜马拉雅为其分布的中心;大血藤属至少已经知道分布于越南北部,这些属严格地应该称为中国半特有属。
|
|
大围山种子植物区系具有如下特征:1)科属种组成丰富,属内种系相对贫乏,分化程度较弱;2)热带分布科属占绝对优势,热带亚洲植物区系特点明显;3)植物区系的古老性明显,古老的特有属丰富;4)大围山是多种区系成分的交汇地带,其植物区系与东亚区系,特别是与北部湾植物区系的联系较为紧密。
大围山虽然地处北回归线附近,理论上应受北回归线副热带高压的下降气流的影响,本应成为干热少雨区域(如西亚、非洲、南美洲同纬度范围的干旱地区),但实际上却是受来自太平洋南中国海夹带充沛暖湿气团的东南热带季风所控制,而形成同纬度罕见的湿润型的北热带-南亚热带气候,发育了中国大陆湿热性质最强的雨林类型——湿润雨林,也孕育了丰富的植物物种多样性,其保护价值是不言而喻的。大围山在中国植物区系分区中属于古热带植物区北缘的马来西亚森林植物亚区,北部湾地区,这一地区较西部成陆早,属滨太平洋区域,自中生代晚三叠纪“印支运动”以来,就没有经历大的变动,尤其是新生代第三纪以来,气候较为暖湿,马来西亚雨林要素得以伸入和发展,并迅速繁衍起来(周庭儒等, 1984),地史上受第四纪冰期恶劣气候影响较小,为许多古老的植物提供了良好的生存环境。这种特殊的地质历史和地理环境,对森林植物区系的发生和发展带来了深刻影响。因而,大围山为著名的第三纪古热带区系发源地的一部分,保留了十分古老的热带、亚热带区系。古老的热带起源区系和进化的亚热带区系融合,类群分化,形成以古老热带区系成分为主、种类组成丰富、多种区系成分并存、物种的水平替代和垂直分布变化十分显著的植物区系特征。
郭辉军, 龙春林. 1998. 云南的生物多样性. 昆明: 云南科技出版社, 38-56.
|
姜汉侨. 1980. 云南植被的分布特点. 云南植物研究, 2(1): 23-24. |
蒋有绪, 王伯荪, 臧润国, 等. 2002. 海南岛热带林生物多样性及其形成机制. 北京: 科学出版社, 111-113.
|
李锡文, 李捷. 1992. 从滇产东亚属的分布论述"田中线"的真实性和意义. 云南植物研究, 14(1): 1-12. |
李锡文. 1995. 云南热带种子植物区系. 云南植物研究, 17(2): 115-128. |
李锡文. 1996. 中国种子植物区系统计分析. 云南植物研究, 18(4): 363-384. |
李延辉, 裴盛基, 许再富, 等. 1996. 西双版纳高等植物名录. 2版.昆明: 云南民族出版社.
|
路安民主编.1999.种子植物科属地理.北京: 科学出版社, 456-457
|
税玉民主编.2003.滇东南红河地区的种子植物.昆明: 云南科技出版社
|
王荷生. 1992. 植物区系地理. 北京: 科学出版社.
|
吴征镒, 朱彦丞. 1987. 云南植被. 北京: 科学出版社, 34-36.
|
吴征镒, 周浙昆, 李德铢, 等. 2003. 世界种子植物科的分布区类型系统. 云南植物研究, 25(3): 245-257. DOI:10.3969/j.issn.2095-0845.2003.03.001 |
吴征镒. 1991. 中国种子植物属的分布区类型. .云南植物研究, (增刊, Ⅳ): 1-139. |
西南林学院, 云南省林业厅, 临沧地区行政公署林业局, 等. 2004. 中国南滚河国家级自然保护区. 昆明: 云南科技出版社, 4-5.
|
自然保护区.昆明: 云南科技出版社, 4-5
|
应俊生, 张玉龙著. 1994. 中国种子植物特有属. 北京: 科学出版社, 12-15.
|
周庭儒, 任森厚. 1984. 中国自然地理—古地理. 北京: 科学出版社.
|