 2006, Vol. 42
2006, Vol. 42文章信息
- 李意德, 方洪, 罗文, 陈焕强, 蒋忠亮.
- Li Yide, Fang Hong, Luo Wen, Chen Huanqiang, Jiang Zhongliang.
- 海南尖峰岭国家级保护区青皮林资源与乔木层群落学特征
- The Resource and Community Characteristics of Vatica mangachapoi Forest in Jianfengling National Nature Reserve, Hainan Island
- 林业科学, 2006, 42(1): 1-6.
- Scientia Silvae Sinicae, 2006, 42(1): 1-6.
-
文章历史
- 收稿日期:2004-02-10
-
作者相关文章


2. 海南尖峰岭国家级自然保护区 乐东 572542
2. Jianfengling National Nature Reserve of Hainan Province Ledong 572542
青皮(Vatica mangachapoi)属龙脑香科(Dipterocarpaceae),为典型热带雨林树种,是我国海南岛中南部地区常见树种,尤以尖峰岭、霸王岭、猕猴岭等地集中,是热带常绿季雨林(蒋有绪等,1991)或称热带低地雨林(胡玉佳等,1992)的建群种。青皮材质坚硬,结构细致,心材极耐腐蚀,是珍贵的阔叶硬材树种。人口增长、不合理采伐以及对青皮专性利用等原因导致了青皮林资源数量急剧下降(李意德,19 95;曾庆波等,1997;李意德等,2002a)。为保护好这一珍贵的森林资源,195 6年划定尖峰岭的五分区和三分区为禁伐区,其中三分区以青皮林为主,总面积381 hm2,是海南岛目前保存最好的青皮林。为了摸清这一珍贵森林资源,了解其种群和群落学特性,于2002年底对该区的青皮林进行了系统的资源调查,为保护、恢复、发展和有效管理青皮林资源提供了科学依据。
1 试验区概况尖峰岭林区位于海南岛乐东和东方两县交界处(18°36′—18°52′N,108°52′—109°5 ′E),面积47 227 hm2,低纬度热带岛屿季风气候,年平均气温24.5 ℃,≥10 ℃的年积温约9 000 ℃,最冷月平均气温19.4 ℃,最热月平均气温27.3 ℃,干湿两季明显,5—10月为雨季,11月至翌年4月为旱季。尖峰岭地区从沿海至最高海拔1 412.5 m处,形成了明显的气候、土壤和植被垂直带谱系列,主要植被类型有滨海刺灌丛、稀树草原(或稀树灌丛)、热带半落叶季雨林、热带常绿季雨林、热带北缘沟谷雨林、热带山地雨林、山地常绿林和山顶苔藓矮林等,其中地带性植被类型为热带常绿季雨林(黄全等,1986;蒋有绪等,1991)。
尖峰岭国家级自然保护区位于尖峰岭林区腹地,面积20 170 hm2,是我国保存面积较大、结构和功能最好的热带原始林区之一。三分区位于保护区西北部,为核心区的一部分,海拔250~850 m,主要森林类型为常绿季雨林,分布在海拔300~700 m的山体中部,由于海拔升高,降雨量比分布海拔低的热带半落叶季雨林有较大幅度的增加,空气相对湿度也相应增大,虽有短暂的旱季,但干旱程度较轻。群落外貌全年常绿,以龙脑香科树种青皮占优势,属于湿润低地热带雨林类型(胡玉佳等,1992)。除青皮外,其他常见种为:细子龙(Amesiodendron chinensis)、野生荔枝(Litchi chinensis var. euspontanea)、海南紫荆木(Madhuca hainanensis)、盘壳栎(Cyclobalanopsis patelliformis)、倒卵阿丁枫(Altingia obovata)、白茶(Koilodepas hainanensis)等。下木层以棕榈科(Palmae)植物为多,如穗花轴榈(Licuala fordiana)、红藤(Daemonorops margaritae)、钩叶藤(Plectocomia microstachys)等。草本层植物稀少,仅见海南砂仁(Amomum longiligulare)、山姜属(Aplinia spp.)植物、单叶新月蕨(Pronehrium simplex)等。层间植物以大型木质藤本为主,如买麻藤(Gnetum montanum)、过江龙(Entada phaseoloides)、黄连藤(Arcangelisia loureiri)、鸡血藤(Millettia reticulata)等,附生植物有崖姜蕨(Pseudodryn aria coronans)、鸟巢蕨(Neottopteris nidus)等,但数量较少(蒋有绪等,1991;黄全等,1986)。
2 研究方法采用典型取样法,从海拔约250 m处的热带半落叶季雨林至海拔约850 m的热带山地雨林,中间包括热带常绿季雨林,沿海拔梯度在林分结构完好的地段设置23个样方(海拔在1: 10 000地图上反查),每个样方面积20 m×30 m,总调查面积1.38 hm2。对样方内胸径≥12 cm的所有乔木树种记载种名、胸径、树高、活枝下高等林分因子。
计算平均胸径:先求算样方中各树木的胸高断面积,然后累加,再求平均断面积,最后反求出平均胸径。
求算蓄积量:采用材积模型法。热带森林组成种类繁多,又没有明显的优势种群(黄全等,1986;1988;蒋有绪等,1991;曾庆波等,1997;李意德,1997),因此对所有种群都建立相应的材积模型是困难的。根据前人的研究资料,尖峰岭地区的树木按照生长类型可分为速生型、中生型和慢生型3大类,速生型和慢生型树种所占的比例较小,大约10%,中生型比例最大,为了简化计算,我们采用了中生型的树木材积混合模型来计算单株材积,然后累加得出各样方的蓄积量参数。中生型树木的材积模型(蒋有绪等,1991)为V=1.809 48×10-4D2.417 860 94。式中V为单株材积(m3),D为树木胸径(cm)。
种群重要值计算:以三分区的森林为总体单元,以23个样方资料为样本单元,依据取样数据,计算各种群的相对密度、相对优势度和相对频度,累加这3个指标后即为种群重要值(Cox, 1979),所有种群重要值之和为300。
采用Excel 2003(Microsoft Corporation)和Statistica 8.0(StatSoft Inc.)等软件进行数据分析和制图。
3 结果分析 3.1 林分因子与森林蓄积量林分因子包括各样方的物种数、植株数量、平均胸径、平均树高、种群蓄积量等。各样地的林分因子详见表 1。表 1中未列出平均树高这一指标,因为在热带林中估测的树高误差较大。
|
|
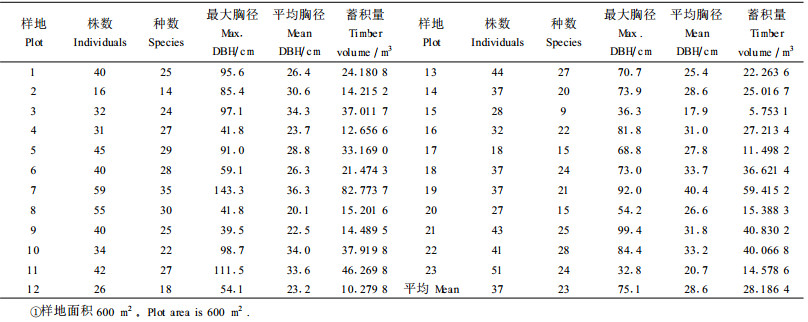
由表 1可以看出,23个样方中胸径≥12 cm的乔木树种每600 m2平均37株,最多59株,最少16株;种群数量平均23种,最多35种,最少只有9种;平均胸径为28.6 cm,最大平均胸径为40.4 cm,最小平均胸径为17.9 cm;最大植株的胸径平均值为75.1 cm,最大胸径143.3 cm,最小胸径32.8 cm,有15个样方出现了胸径≥70 cm的最大植株,占样方总数的65%,说明三分区的森林资源目前仍保存地较为完好,具有较强的原生性。
23个样方的平均蓄积量为28.186 4 m3,即单位面积蓄积量为469.773 m3·hm-2,作为森林的平均蓄积量,这一指标是相当高的,例如1958年尖峰岭森林基本上未受采伐和其他活动影响时调查所得蓄积量为280~390 m3·hm-2(蒋有绪等,1991)。从最高蓄积量水平来看,600 m2中竟有高达82.773 7 m3的样方(7号),其单位面积蓄积量达1 380 m3·hm-2,这样高的蓄积量,与典型取样(如在1株或多株大树的范围内选取)有较大的关系,在该样方出现了1株胸径达143 cm的大叶榕(Ficus viries var. sublanceolata)和2株胸径分别为85和99 cm的盘壳栎。但15号样方蓄积量只有5.753 1 m3,单位面积蓄积量仅96 m3·hm-2,不仅大大低于本次调查的平均水平,而且低于半落叶季雨林平均值145 m3·hm-2(蒋有绪等,19 91),主要是由于树木密度较大而平均胸径较小的缘故。
在组成树种中,青皮的蓄积量最大,达28.650 7 m3·hm-2,占总蓄积量的6.10%,反映了青皮相对占优势;其伴生种类盘壳栎为27.810 5 m3·hm-2,占5.92%;野生荔枝24.656 5 m3·hm-2,占5.25%;大叶榕24.613 8 m3·hm-2,占5.24%;橄榄(Canarium album)17.224 2 m3· hm-2,占3.67%。这5个优势种类占总蓄积量的26.17%。而其他种类的蓄积量较小,如热带常绿季雨林表征种细子龙(蒋有绪等,1991;曾庆波等,1997;李意德等,2002b)的蓄积量只有12.773 6 m3·hm-2,仅占2.72%,毛果稠(Lithoca rpus pseudovestitus)的蓄积量和所占比例分别为12.032 8 m3·hm-2和2.56%,所占比例为2%~3%的有7个种,1%~2%的有14个种,绝大多数种(约占总种数的92%)的蓄积量比例只占1%以下,充分说明了常绿季雨林种类构成的多样性导致了蓄积量构成的复杂性。
3.2 种群重要值分析23个样方中,共出现了胸径≥12 cm的乔木物种314个,占尖峰岭地区已知乔灌木种类(1 000余种)的30%以上(曾庆波等,1995),物种多样性水平很高。为更好地探讨各主要物种在群落中的地位和作用,将三分区的森林乔木层作为一个植被总体单元,用23个样方资料进行重要值等群落学特征分析,重要值≥2.0的32个种群见表 2,其他种群的重要值均小于2,属于偶见种群。
|
|
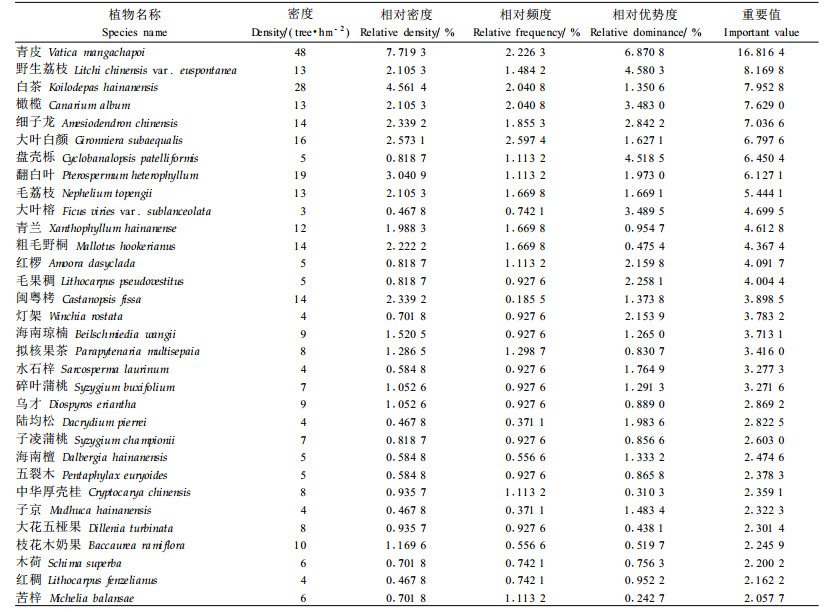
从表 2可看出,常绿季雨林的表征种如青皮、野生荔枝、白茶、橄榄和细子龙构成了该区的优势种群,其他次优势种群包括大叶榕、红椤、海南琼楠、乌才、枝花木奶果等;翻白叶、海南檀种等则主要见于常绿季雨林的分布下限和半落叶季雨林中;而大叶白颜、盘壳栎、毛荔枝、青兰、粗毛野桐、毛果稠、灯架、拟核果茶、水石梓、碎叶蒲桃、子凌蒲桃、中华厚壳桂、子京、大花五桠果、木荷、红稠、苦梓等在常绿季雨林和山地雨林为共有种类;陆均松、五裂木等主要见于海拔较高的山地雨林;闽粤栲是海拔700~900 m采伐迹地上的先锋种(黄全等,1986),本次调查仅在海拔800~900 m的山地雨林采伐迹地更新林中出现。
分析结果表明,青皮在三分区森林群落中所起的作用最大,大约占5.6%的群落份额(即重要值百分比为16.8/3=5.6),野生荔枝、白茶、橄榄和细子龙等常绿季雨林表征种类分别占了2.3%~2.7%的群落份额(表 2),与前人对一片典型青皮林采用点-四分法的研究结果相比(黄全等,1986;蒋有绪等,1991),由于本次调查取样范围较大,异质性高(涵盖了部分半落叶季雨林和山地雨林类型),因此这5个表征种类的重要值都有不同程度的降低,排序也不尽相同,如在点-四分法取样中,最优势种类为白茶(重要值百分比为14),其次为橄榄(11.1)和细子龙(11.1),青皮排在第4位(10.4),但这4个种的差别不大,野生荔枝则排位靠后(7.0)。总之,不管其取样方法和范围如何,在尖峰岭常绿季雨林中,青皮、白茶、野生荔枝、橄榄和细子龙5个种类可认为是该植被类型的表征种,尽管它们的重要值大小会有一些差异,其排序位置也可能会发生变化,但是群落的本质不会改变,除非有人为的干扰和严重的自然干扰发生。在蓄积量排序中处于第2位的盘壳栎和处于第4位的大叶榕,由于相对密度和相对频度很低,尽管相对优势度较高,但其重要值仅排第7和10位,表明这2个种在群落中所起的作用是局部的而不是全局的。
3.3 样方的排序三分区17、20和21林班的海拔为250~850 m,包含了半落叶季雨林、常绿季雨林和山地雨林3个主要植被垂直类型,半落叶季雨林和山地雨林2个植被带的范围较小,分别为50~100和150~200 m,主要为常绿季雨林及其向两侧伸延的交错带(ecotone)类型。将这23个样方进行聚类分析,种群组成比例为变量,可形成3个主要样方组(图 1):组Ⅰ包括样方8、12、13、17和20,以白茶、翻白叶等占优势,间或出现青皮、细子龙等种类,显然这几个样方属于常绿季雨林向半落叶季雨林过渡的类型,海拔250~450 m;组Ⅱ包括样方5、7、9、10、11、14、16、18和19,其种群以青皮为主,间有白茶、细子龙、野生荔枝、橄榄等种类,是典型的常绿季雨林类型,海拔300~650 m;组Ⅲ包括样方1、2、3、4、6、21和22,种群组成以大叶白颜、粗毛野桐、中华厚壳桂、毛荔枝等为主,部分样方含有青皮、细子龙等种类(如1—4号样方),基本上属于常绿季雨林向山地雨林伸延的过渡类型,海拔600~800 m,只有6、21和22号样方为不含青皮的典型山地雨林。另外,15和23号样方在聚类分析中成为孤立样方,因为15号样方以耐旱的黄牛木(Cratoxylon ligustrinum)为主,属典型的半落叶季雨林,而23号则为本次调查海拔最高(约850 m)的样方,并且以闽粤栲为主,显然它是由山地雨林采伐后更新起来的先锋群落(黄全等,1988)。

|
图 1 尖峰岭三分区青皮林23个样方聚类树状图 Fig. 1 Tree-diagram of 3 plots based on clustering analyse 距离系数为欧几里得距离的平方,采用韦式方法聚类 Using squared Euclidean distances and Ward's clustering method |
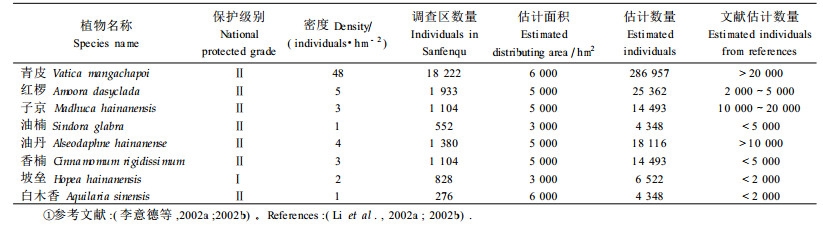
参照国家重点保护植物名录(第一批,1999年)对本次调查出现的物种进行统计,共有8种国家重点保护植物(表 3),其中坡垒(Hopea hainanensis)为国家一级重点保护植物;国家二级重点保护植物有7种。在调查区内(381 hm2),估计青皮的数量达18 000株以上,其他7个物种的种群数量为276~1 933株。依据这8个物种的生态生物学特性和在尖峰岭地区的适生分布范围,估算各种群数量:青皮286 000株以上,红椤25 000株以上,均高于文献记载的估计值;香楠(Cinnamomum rigidissimum)、坡垒、白木香(Aquilaria sinensis)、子京、油楠(Sindora glabra)和油丹(Alseodaphne hainanense)等与文献估计数量相差不大。
|
|
鉴于青皮为常绿季雨林的优势种,对其进行种群分析,有助于对该物种的保护和持续利用。样方中共记载了青皮66株,密度为48株·hm-2,最大胸径60 cm。我们以青皮的径级分布来探讨青皮的种群动态,因为树木胸径是年龄的函数(蒋有绪等,1991;Li et al., 1997),利用径级对应植株数量可看出该物种的结构是否合理,并能判断种群发展动态,其结果见图 2。可以看出,青皮种群小径级植株数最多,如径级12~1 6 cm的株数为15株,而大径级植株仅3~5株,并且这种下降趋势是渐进的,其种群结构呈反J曲线(reverse J curve),这是一种良好的结构类型(Li et al., 1997),表明尖峰岭三分区的青皮种群为成熟的增长型种群,其未来发展是正常的。

|
图 2 植体胸径级分布 Fig. 2 Distribution of DBH grade |
三分区分布着目前海南岛保存最好的天然青皮林,自1956年成立禁伐区以来基本上未受到干扰破坏,由于该区靠近中沙、江边等居民聚居地而偶有盗伐青皮等珍贵木材的现象发生,但这种小范围和极低强度的干扰还不足以导致森林生态系统发生质的退化,本次调查结果可充分说明这一点。从群落结构、种类组成、林木径级(特别是优势种青皮的径级分布)、森林蓄积量等指标来看,均表明该区森林的原生性非常强。
该青皮林属于我国热带季风气候条件下的典型地带性植被类型——热带常绿季雨林,在尖峰岭地区的最佳分布海拔300~650 m处,向下过渡到热带半落叶季雨林,偶有植株分布到海拔200 m处;向上过渡到热带山地雨林,个别植株见于海拔750 m处。由于共有种的存在,各植被类型之间没有明确的界线,有100~200 m的生态交错区。
青皮是热带常绿季雨林生态系统的关键种(key stone species),对生态系统的健康与持续发展将起到非常重要的作用。要保护好森林,首先要保护好关键种,因为一个关键种的兴衰,将直接导致10~20个相关物种的兴衰,而这些物种又可导致其他相关物种的兴衰(Mc Neely,1990;张知彬,1994)。因此保护好青皮物种,是保护好这片森林及其他国家重点保护动植物资源的关键。根据青皮种群组成结构较优、种群数量较大(仅三分区就超过了18 000株)的现状,加之目前该区域的保护措施比较到位,青皮在未来的较长时间内将处于正常的发展态势。三分区目前没有居民居住,区内的人为影响几乎不存在,但该区外围一些村镇的居民对青皮有专性利用的习惯(李意德,1995),胸径20~30 cm、树干通直的青皮是他们做建筑、家具、农具的良材(从图 2可看出,胸径20~30 cm的植株相对较少一些,可能与此有关),偶有盗伐青皮行为。为此,应当在三分区外围设立固定的护林点,以有效地保护这片珍贵的森林资源。
胡玉佳, 李玉杏. 1992. 海南热带雨林. 广州: 广东高教出版社.
|
黄全, 李意德, 郑德璋, 等. 1986. 海南岛尖峰岭地区热带植被生态系列的研究. 植物生态学与地植物学学报, 10(2): 90-105. |
黄全, 李意德. 1988. 海南岛尖峰岭热带山地雨林采伐迹地更新群落的初步分析. 植物生态学与地植物学学报, 12(10): 12-22. |
蒋有绪, 卢俊培. 1991. 中国海南岛尖峰岭热带林生态系统. 北京: 科学出版社.
|
李意德, 林玉梅, 郭宁, 等. 2002a. 海南尖峰岭地区珍稀濒危植物及其丰富度. 热带林业, 30(4): 35-39. |
李意德, 陈步峰, 周光益, 等. 2002b. 中国海南岛热带森林及其生物多样性保护研究. 北京: 中国林业出版社.
|
李意德. 1997. 海南岛尖峰岭热带山地雨林的群落结构特征. 热带亚热带植物学报, 5(1): 18-26. |
李意德. 1995. 海南岛热带森林的变迁及生物多样性的保护对策. 林业科学研究, 8(4): 455-461. DOI:10.3321/j.issn:1001-1498.1995.04.003 |
曾庆波, 李意德, 陈步峰, 等. 1995. 海南岛尖峰岭地区生物物种名录. 北京: 中国林业出版社.
|
曾庆波, 李意德, 陈步峰, 等. 1997. 热带森林生态系统研究与管理. 北京: 中国林业出版社.
|
张知彬. 1994.生物多样性保护的若干理论基础//中国科学院生物多样性委员会编.生物多样性的原理与方法.北京: 中国科学技术出版社, 36-54
|
Cox G W. 1979.普通生态学实验手册.蒋有绪, 译.北京: 科学出版社
|
Mc Neely J A. 1990.保护世界的生物多样性.薛达元, 等译.北京: 中国环境科学出版社
|
Li Yide, Comiskey J, Dallmeier F. 1997. Structure and composition of tropical mountain rain forest at Jianfengling Nature Reserve, Hainan Island, P. R. China//Dallmeier F. Forest biodiversity in Europe, Australia, and Africa: Res earch, Monitoring and Modeling. USA: The Parthenon Press, 551-562
|