 2005, Vol. 41
2005, Vol. 41文章信息
- 李景文, 聂绍荃, 安滨河.
- Li Jingwen, Nie Shaoquan, An Binhe.
- 东北东部林区次生林主要阔叶树种的萌芽更新规律
- Stump Sprouting of the Main Broad-Leaved Tree Species of Secondary Forest in Eastern Area of Northeast China
- 林业科学, 2005, 41(6): 72-77.
- Scientia Silvae Sinicae, 2005, 41(6): 72-77.
-
文章历史
- 收稿日期:2003-06-16
-
作者相关文章


2. 东北林业大学 哈尔滨 150040;
3. 山东省泰安市泰山风景管理委员会 泰安 271000
2. Northeast Forestry University Harbin 150040;
3. Taishan Landscape Management Council, Shandong Province Tai'an 271000
无性更新是利用留存在迹地上的树桩(伐桩)和根系上的休眠芽或不定芽形成新的植株,以达到更新的目的,萌芽更新与根蘖更新均属无性繁殖方式。由于无性更新省略采种、育苗、植苗等一系列过程,因而是所有更新方法中最为省时、省工的(叶镜中等,1989)。萌芽更新的物质基础是伐桩上的休眠芽和不定芽。休眠芽由原生分生组织形成,不定芽由次生分生组织形成。几乎所有的阔叶树都能由休眠芽生长萌条,休眠芽保持萌发的能力因树种生物学特性的不同而差别较大(叶镜中等,1989)。
萌芽更新在林业上意义很大,尤其在我国广大的次生林地区占有重要地位,主要体现在择伐与皆伐迹地的森林恢复和演替以及苗木培育等具有一定的作用(河北省林业实验场,1958;叶镜中等,1989;于明坚,1999;喻理飞等,2002),特别是对于强度择伐的天然林恢复及人工林天然化经营有较大意义(李景文等,2000)。有关树木无性更新的研究较少,且这些研究主要集中于单个树种,如研究较多的栎树(Quercus spp.)(李克志,1958;Johnson,1977;David et al., 1998;张革等,1999)、杉木(Cunninghamia lanceolata)(马祥庆等,1994;高健等,1995;林武星等,1996;何宗明等,2000;王传佳,2000)、山杨(Populus davidiana)(张正昆等,1984)和刺槐(Robinia pseudoacacia)(王进鑫等,1994;董金伟等,2001)等少数几个树种;研究的内容多侧重于萌芽数量、生长情况、不同经营方式对萌芽更新的影响等。对于萌芽更新规律的研究也有报道(Johnson,1975;叶镜中等,1989;王进鑫等,1994;李景文等,2000);但这些研究缺乏对萌芽动态以及“萌代主”等内容的研究。本文探讨东北东部林区次生林主要阔叶树种伐桩萌芽产生规律、萌芽发展动态和“萌代主”等问题,为退耕还林、次生林的改造和重建以及人工林的天然化经营提供理论依据。
1 研究地概况研究地点位于黑龙江省依兰县西南部的先锋林场,地理坐标为129°27′—129°44′E,45°50′—45°03′N。该区海拔250~500 m,平均气温3 ℃,大于10 ℃的年积温2 580 ℃,无霜期130 d左右,夏季多雨,降水量在323.9 mm,多集中在7、8月份;年平均蒸发量1 340.7 mm;土壤是以暗棕壤为主。
试验地设于先锋林场的第57林班第4小班,坡度5~10°;光照及排水条件良好;土壤pH值为6.8;植被为经过择伐的针阔混交林(1997年冬季择伐),郁闭度0.3左右,主要乔木包括红松(Pinus koraiensis)、水曲柳(Fraxinus mandshurica)、黄檗(Phellodendron amurense)、胡桃楸(Juglans mandshurica)、榆树(Ulmus spp.)、槭树(Acer spp.)等,灌木与木质藤本有狗枣猕猴桃(Actinidia kolomikta)、鸡树条荚迷(Viburnum sargenti)、东北山梅花(Philadelphus schrenkii)等;草本植物包括木贼(Equisetum hyemale)、蚊子草(Filipendula palmata)等。
2 研究方法 2.1 野外调查以往的研究表明,影响树木萌芽更新规律的主要因素是采伐时间、立地条件、伐桩年龄、萌芽部位(李克志,1958;叶镜中等,1989),同时伐桩径和高度对于萌芽数量也有一定作用(黄世能,1990;张加正等,1994;李景文等,2000)。本研究选取依兰县先锋林场1997年择伐林地设置固定样地。样地择伐前为次生红松针阔混交林,面积2.5 hm2。择伐后立地条件、择伐强度、植被类型均较为一致。分别于1998和1999年对样地阔叶树伐桩的年龄、径与高度,萌生条产生的部位(伐桩的基部为距地面距离≤10 cm;中部为距地面距离10~20 cm。实际调查中,伐桩的高度差别较大。该区分标准主要考虑多数伐桩高度值和实际经营中应用方便以及统计的一致性,而20 cm以上的部位无论伐桩的高度多大都定义为伐桩的上部)、萌生条的数量(包括总数、活枝数、死亡数)、高生长(平均高)以及基径进行了调查。在萌代主动态的调查中,采用“时空替代”的方法,对择伐时间相对较长的林地伐桩萌芽更新的萌生树的数量、高度和径生长的情况进行了调查。
2.2 数据的统计和分析在对伐桩萌芽更新调查中,共测量伐桩304个,对于高度大于80 cm的伐桩,在实际经营中没有意义,在分析中略去,实际分析了290个伐桩。在萌芽数量与伐桩径大小分析中,主要以5 cm为伐桩径级区分单位(0~5 cm为1级,6~10 cm为2级,10~15 cm为3级,依次类推,而槭树的伐桩数量较少,以10 cm为径级区分单位)分别统计各径级内萌生枝的平均数量。同时,在分析萌芽数量与伐桩高的关系中,主要也以5 cm为区分高度级,统计各阔叶树种在各高度范围内伐桩所产生萌生枝的平均数量;并对萌芽与伐桩径和高度之间变化趋势进行了模拟。考虑到山槐(Maackia amurensis)、紫椴(Tilia amurensis)、稠李(Padus asiatica)等树种伐桩数量太少而未做分析;同时,在调查中发现槭树和榆树等属内的各树种的伐桩萌芽发生规律具有一致性,故归槭树类和榆树类进行统计。对不同树种2年生萌芽保存率,数量特征以及生长情况进行了对比。在萌代主动态的分析中,采用“时空互代”的方法,分析了萌代主萌生树数量和生长状况。
3 研究结果 3.1 主要阔叶树种萌芽更新的一般规律在环境条件相同的情况下,伐桩产生萌芽数量取决于自身特性。通过对样地内萌芽更新的调查,经分析得出样地内的水曲柳、胡桃楸、黄檗、榆树、槭树等阔叶树种的伐桩都有萌芽产生(表 1)。
|
|
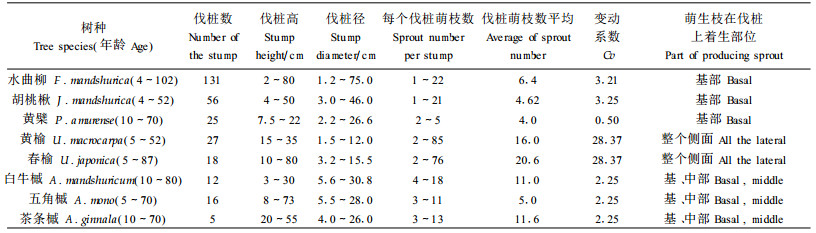
伐桩产生萌芽的数量因树种不同而存在较大的差别,而且同一树种不同直径和高度的伐桩所产生萌芽数量也不同。从表 1看出,榆树的变动系数最大,高达28.37,其他树种变动系数差别不大,一般在2~4之间;而黄檗变动较小,仅为0.5。各树种的伐桩萌芽产生的部位不同:三大硬阔的萌芽主要发生在伐桩的基部,而榆树的萌芽在整个伐桩的侧面都有分布,且上部较多;而其他树种一般在伐桩的中下部。
3.2 伐桩产生萌芽数量的规律 3.2.1 萌芽数量与伐桩径的关系在立地条件相同的情况下,萌芽数量主要取决于伐桩年龄、直径和高度等因素(李克志,1958;叶镜中等,1989;黄世能,1990)。伐桩的直径和年龄具有相关性,以往有关萌芽更新规律的研究多采用伐桩直径(Johnson,1975;1977;MacDonald et al., 1983;黄世能,1990;张加正等,1994;David et al., 1998;李景文等,2000)。另外,利用直径得出的数据在森林经营中易于操作。因此,本文将结合伐桩径和高度与萌芽数量的关系探讨萌芽的产生数量规律。
伐桩产生萌芽的数量和高生长与伐桩直径、年龄有关,而且这一关系因树种的不同也存在差别(表 2)。本研究样地内各树种不同径级的伐桩都具有萌芽的能力,产生的萌芽数量随着伐桩直径增加而增大,达到一定伐桩直径后又随着伐桩直径增加而减少。从树种伐桩的萌芽的数量和高生长分析,水曲柳、胡桃楸、黄檗、榆树在径级分别为10~20、15~20、10~ 15、5~20 cm,年龄在20~50、40~50、40~70、10~80年的萌芽能力较强;而槭树小径木萌芽能力较高。同时各树种的伐桩在10~20 cm不但萌生能力很强,而且高生长也具有明显的优势,当年生的萌生条高度都在1 m以上;远远大于各树种当年生的实生苗高生长。
|
|
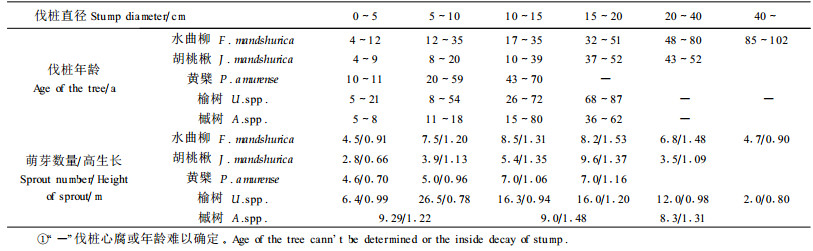
对当年生萌生枝数随伐桩径的变化趋势进行拟合得萌生枝数与伐桩径的关系回归模型(表 3)。各树种伐桩产生的萌芽数与伐桩径大小呈多项式分布,该结果与Johnson(1975)对红槲栎(Quercus rubra)萌芽规律研究结果相近。
|
|

伐桩产生萌芽的数量和伐桩的高度有关。对实测的伐桩进行分析,统计各伐桩高度范围萌生枝的平均值,并对伐桩高度与萌芽的数量关系进行分析(表 4)。
|
|
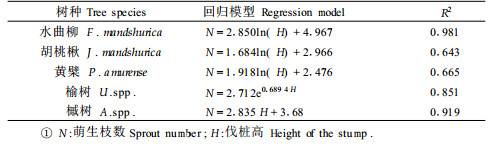
从表 3的分析可以得出,各树种伐桩所产生的萌芽数量与伐桩高度级的关系存在着3类不同的变化趋势:水曲柳、胡桃楸和黄檗的变化趋势基本一致,随伐桩高度的增大,萌芽数量变化较平缓;槭树伐桩产生的萌生枝数量与伐桩高度级的关系表现出直线变化趋势;榆树萌生枝数量随着伐桩高度级的增大而剧增,呈指数变化趋势。结合萌芽数量与伐桩直径的关系分析可见,水曲柳、胡桃楸和黄檗产生萌芽能力主要和伐桩直径相关,而榆树、槭树萌芽能力和伐桩的直径和高度都有关,特别是榆树萌芽的数量和伐桩高度关系更大。
3.2.3 萌芽的数量与高生长状况在萌生枝的高生长方面,各树种表现出的规律与萌芽的数量与伐桩径的关系基本一致,首先随着伐桩径的增大,萌生条的高生长增加(表 2);到伐桩径在20 cm左右,随伐桩径的增大而降低;与伐桩高度的关系不明显。水曲柳、黄檗、胡桃楸当年生萌生枝的高度一般在80~170 cm之间;基径的大小一般为0.5~2.6 cm; 槭树当年生萌生枝高度与三大硬阔差别不大;榆树的当年生萌生枝的高度一般在40~120 cm之间, 基径一般为0.3~0.9 cm。
3.3 伐桩萌芽数量动态与萌代主状况分析为探讨伐桩所产生萌生枝的动态变化,于1999年8月对样地进行了复查,共测得201个活伐桩。分析了不同树种活伐桩的比率、伐桩次年萌生枝数量动态(由于榆树和槭树类存活率很低,伐桩数量少,所以未统计)及高生长状况。
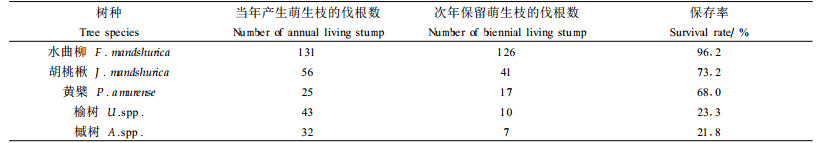
3.3.1 不同树种活伐桩的保存率具有存活萌生枝的伐桩占固定样地内该树种伐桩总数的比率,即保存率。不同树种活伐桩的保存率差别较大(表 5)。其中水曲柳、胡桃楸及黄檗的保存率较高,特别是水曲柳活伐桩的保存率高达96.2%;而榆树和槭树伐桩的存活率略高于20%,明显低于三大硬阔类。而且各树种的保存率在5~10 cm径级内较高。
|
|
伐桩萌生条数量与伐桩径的关系在总体趋势上与第1年基本相同;同时,调查中还发现第2年仍有新的萌生条产生,但数量很少,且发育不良。与第1年不同之处在于萌生条开始死亡,并且伐桩产生的萌生枝越多,死亡的数量也增大(图 1)。

|
图 1 水曲柳、胡桃楸、黄檗的伐桩径与2年生萌生枝数量动态 Fig. 1 Dynamics between the total, living and dead number of biennial sprout and stump diameter of Fraxinus mandshurica, Juglans mandshurica and Phellodendron amurense
水曲柳的伐桩径级1为0~5 cm, 2为5~10 cm, 3为10~15 cm, 4为15~20 cm,5为20~40 cm。 The class of stump diameter for F.mandshurica: 1 is 0~5 cm, 2 is 5~10 cm, 3 is 10~15 cm, 4 is 15~20 cm,5 is 20~40 cm. 胡桃楸伐桩径级1为0~5 cm, 2为5~10 cm, 3为10~15 cm, 4为15~20 cm,5为20~50 cm. The class of stamp diameter for J. mandshurica: 1 is 0~5 cm, 2 is 5~10 cm, 3 is 10~15 cm, 4 is 15~20 cm,5 is 20~50 cm. |
高生长方面,水曲柳、胡桃楸和黄檗2年生的萌生枝高度分别为180~280、190~240和180~250 cm。
3.3.3 萌代主分析萌芽更新的意义关键在于萌生枝能否代替被采伐的主干形成大树,即“萌代主”的问题。本研究中采用“时空互代”,分别调查了较早的采伐林地。调查和分析表明,本文所讨论树种伐桩的萌生条都存在“萌代主”的大树,但所形成大树的数量不确定(表 6)。如水曲柳的伐桩所保留的10年生萌生枝平均3株左右,胡桃楸42年生萌生大树可多达7株。
|
|

在调查中发现榆树和槭树“萌代主”和树种径级以及部位有很大关系。较大径级(20 cm以上)的伐桩,“萌代主”的萌生条主要在伐桩的基部,而随伐桩直径的减小(10~20 cm),“萌代主”的萌生条位置上移;5~10 cm小径木的“萌代主”的萌生条主要在伐桩的上部。
4 讨论把研究地点具有萌芽更新能力的阔叶树种划为3种类型:对数型、直线型、指数型。以往的研究多为单个树种无性更新一般规律及其经营的研究。从各树种萌芽更新的特点可见,三大硬阔类萌芽更新在自然条件下“萌代主”伐桩的保存率和成功率都很高,特别是水曲柳和胡桃楸的保存率更高,利用价值更大;槭树、椴树、榆树的萌芽更新也有一定价值。结合各树种的萌芽能力和活伐桩的保存率的分析结果可以看出5~20 cm伐桩在利用无性更新中是重点经营对象,10~20 cm伐桩所产生的萌芽生长最好。对于如何通过人工控制萌生枝的数量而提高各树种的保存率还有待进一步研究。
“萌代主”问题,要重视在所产生的萌芽中,哪些萌芽是经营中应重点保留的,保留多少为宜。过去一般认为重要的是树干最下部的休眠芽(叶镜中等,1989)。本研究发现,三大硬阔的15 cm以上径级的伐桩萌芽符合上述规律。而其他阔叶树中不同树种、径级和高度的伐桩,萌代主的萌生枝着生的部位不同。如槭树、蒙古栎(Quercus mongolica)、榆树等树种的小径木(10 cm以下)萌代主的萌生树主要分布于伐桩上部,伐桩高度越大、伐桩径越小越明显;同时,三大硬阔中15 cm以下伐桩“萌代主”的萌芽和大径级伐桩相比位置上移。
对于每个伐桩“萌代主”萌芽保存的数量,研究的结果是否完全符合不同树种“萌代主”的规律,还有待于大量数据的证明,但确实表明哪些树种所产生的萌芽可以“萌代主”;同时表明,虽然每个伐桩“萌代主”的萌芽数量差别较大,但从平均数量来看,一般为2~3个;这也可以初步推断在萌芽更新中,每个伐桩应该保留2~3个生长良好的萌芽为宜。“萌代主”萌芽保留的数量过多,萌生的大树的干性不良,即所谓“砍刀形”大树,木材利用价值降低。
群落稳定性和植物的冗余有一定的关系(党承林,1998)。在东北地区阔叶树种的萌芽冗余在维持森林群落的稳定性及其恢复方面有一定的作用。树木的萌芽初期高生长相对于该树种种子更新幼树要快得多,有利于森林快速更新和恢复(David et al., 1998;李景文等,2000),但在森林群落自然演替中作用很小。对于受人为干扰较大的次生林和人工林的改造和恢复中的地位和作用是明显的。我国很多地区目前有大面积的由于人为干扰而形成的次生林、人工纯林,如果根据各树种萌芽的规律,利用萌芽更新可加速植被的恢复和改善次生林树种的结构并提高次生林的生物多样性。在人工林的改造方面,可以利用林内伐桩产生的萌生条,形成人工林和天然树种混交的状态,对于提高人工林稳定性、生态功能和物种多样性都是很好的途径。因此研究萌芽更新的规律和经营对于天然林保护工程所涉及的次生林的改造和快速恢复、人工林的天然化经营等具有一定的现实意义。
党承林. 1998. 植物群落的冗余结构. 生态学报, 18(6): 665-672. |
董金伟, 杜华兵. 2001. 伐桩不同处理对山地刺槐无性更新影响的研究. 山东林业科技, 4: 10-11. |
高健, 刘令峰, 叶镜中. 1995. 伐桩粗度和高度对杉木萌芽更新的影响. 安徽农业大学学报, 22(2): 145-149. |
河北省林业实验场. 1958. 毛白杨留根繁殖法总结报告. 林业科学, (3): 223-230. |
黄世能. 1990. 不同伐桩直径及高度对马占相思萌芽更新影响研究. 林业科学研究, (3): 242-248. |
何宗明, 杨玉盛, 俞新妥. 2000. 二代实生与萌芽杉木生长特点分析. 福建林学院学报, 20(4): 298-301. |
李景文, 刘世英, 王清海, 等. 2000. 三江平原低山丘陵区水曲柳无性更新研究. 植物研究, 20(2): 215-220. |
李克志. 1958. 柞树萌芽林的研究. 林业科学, (3): 231-247. |
林武星, 叶功富, 黄金瑞, 等. 1996. 杉木萌芽更新的原理及技术评述. 福建林业科技, 23(2): 19-23. |
马祥庆, 王容溪, 陈友力, 等. 1994. 不同林地清理方式对杉木萌芽更新的影响研究. 福建林业科技, 21(2): 15-18. |
王传佳. 2000. 杉木萌芽更新研究. 林业科技开发, 14(2): 23-25. |
王进鑫, 余清珠, 刘增文. 1994. 刺槐人工林无性更新萌芽与根蘖发生规律的初步研究. 陕西林业科技, (2): 27-31. |
吴远由. 1995. 杜仲林伐桩萌芽更新初报. 贵州林业科技, 23(3): 51-52. |
喻理飞, 朱守谦, 叶镜中, 等. 2002. 退化喀斯特森林自然恢复过程中群落动态研究. 林业科学, 38(1): 2-7. |
于明坚. 1999. 青冈长绿阔叶林群落动态研究. 林业科学, 35(6): 42-51. |
叶镜中, 孙多. 1989. 森林经营学. 北京: 中国林业出版社, 19-27.
|
张革, 宋修田, 徐斌. 1999. 辽东山区柞树成、过熟林萌芽更新问题探讨. 辽宁林业科技, (6): 24-25. |
张加正, 黄松秋, 陈伽. 1994. 不同直径伐桩对赤桉萌芽更新影响的初步研究. 浙江林业科技, 14(4): 35-37. |
张正昆, 戴洪才. 1984. 山杨无性更新的研究. 北京林学院学报, (3): 10-18. |
David R Larson, Paul. S. 1998. Linking the ecology of natural oak regeneration to silviculture. Forest Ecology and Management, 106: 1-7. DOI:10.1016/S0378-1127(97)00233-8 |
Johnson P S. 1975. Growth and structural development of red oak sprout stumps. For Sci, 21(4): 413-418. |
Johnson P S. 1977. Predicting oak stump sprouting and sprout development in the Missouri Ozarks. Res. Pap., NC-149, USDA, Forest Service, 11
|
MacDonald J E, Powell G R. 1983. Relationships between stump sprouting and parent-tree diameter in sugar maple in the 1st year following clear-cutting. Can J For Res, 13: 390-394. DOI:10.1139/x83-058 |