 2005, Vol. 41
2005, Vol. 41文章信息
- 季孔庶, 樊民亮, 徐立安.
- Ji Kongshu, Fan Minliang, Xu Li'an.
- 马尾松无性系种子园半同胞子代变异分析和家系选择
- Variation Analysis and Plus Family Selection on Half-Sib Progenies fromClonal Seed Orchard of Pinus massoniana
- 林业科学, 2005, 41(6): 43-49.
- Scientia Silvae Sinicae, 2005, 41(6): 43-49.
-
文章历史
- 收稿日期:2004-02-04
-
作者相关文章


2. 浙江省林业种苗管理总站 杭州 310020
2. Zhejiang Forest Seed and Seedling Administration Hangzhou 310020
子代测定是评价亲本材料遗传价值与估算遗传参数的一条有效途径。我国从“六五”开始, 将主要用材树种的遗传改良列入国家攻关项目, 优树选择和大规模的种子园营建工作已得到广泛开展。在此过程中, 优树的子代测定工作开展得较为及时, 并对评价优树遗传品质的优劣起到了积极作用。而种子园内母树的子代测定, 由于受到开花结实问题的制约而相对滞后。如我国分布最广的针叶树种——马尾松(Pinus massoniana), 优树子代测定在生长量和材质指标方面开展了相关研究(全国马尾松种子园课题协作组, 1990; 周志春等, 1994; 郑仁华等, 1998; 龙光生等, 2002), 为剔除表型选优过程中入选的遗传型并不是优良的“优株”起到了有效作用。马尾松种子园从20世纪70年代末开始营建, 目前已近1 000 hm2(季孔庶, 2001), 从种子园的现状来看, 由于建园材料来自未经遗传测定的表型选择优树, 加上初期栽植密度较大, 因此去劣疏伐问题已成为制约已建种子园种子产量的重要因素。而马尾松种子园的早期疏伐, 只能根据优树子代测定的结果来开展, 原因是马尾松种子园至少在建园5、6年之后才开花, 不可能直接利用园内母树的子代材料来指导疏伐。毕竟优树子代与种子园母树子代相比两者的父本来源不一样, 因此借鉴优树子代测定结果来指导种子园的疏伐存在着一定误差。为有效提高马尾松种子园的种子产量, 最大限度地挖掘遗传潜力, 开展种子园的子代测定和再选择工作已相当迫切。本研究针对福建省漳平五一林场马尾松种子园自由授粉子代材料, 开展生长、形质和材性性状的遗传分析, 对半同胞家系进行评选, 为种子园去劣疏伐提供依据, 并为营建高世代种子园和进一步提高马尾松制浆造纸材改良的遗传增益服务。
1 材料与方法 1.1 种子园及子代测定概况马尾松无性系种子园位于福建省漳平五一林场桂东工区, 东经117°35′, 北纬25°15′; 年均温超过20 ℃, 一月份均温超过10 ℃, 年降雨1 481~1 438 mm, 无霜期310~320 d, 海拔400~500 m, 主坡向为东南阳坡和半阳坡, 土壤肥力中等; 以天然阔叶林、杉木种子园以及阔叶树种人工林为隔离带。全园共计218个无性系, 小区配置15~25个无性系, 总面积20 hm2, 为福建省1980年的选优材料, 接穗来源于福建省官庄林场采穗圃。于1985年定植砧木, 1987年嫁接建成, 初植密度625株·hm-2, 保存率为82%。1991年开始开花结实, 1992年普遍开花结实, 1994—1995年产量最高, 其后每年按原产量15%~35%减少。1997年底在无子代测定数据的情况下, 只得根据种子园内无性系的表型实施了一次疏伐, 强度为0.4, 郁闭度从0.7~0.8降至0.4~0.5。据1999年的测定结果(陈敬德, 2000), 疏伐后的单株球果数比未疏伐前提高了1.3倍。目前该种子园的郁闭度又已接近0.8, 还需去劣疏伐。
1994年采集种子园自由授粉种子, 1995年春季育苗, 并于同年秋季在梅水坑工区营造子代测定林, 参试家系共120个(其中含一个对照CK), CK为福建省漳平当地一般林分的马尾松母树后代, 子代测定林的栽培环境与种子园相似。所有子代测定林试验设计采用完全随机区组, 4株小区, 8次重复。
1.2 方法 1.2.1 调查项目和方法树高、胸径、冠幅、通直度采用常规方法调查, 通直度按优、良、中、差分1~4级, 1级为最好, 最差的为4级[通直度处理时做SQRT(x+1)转换]; 单株材积按原林业部颁布的标准公式计算:V=0.000 062 341 803 D1.856 149 7 H0.956 849 2(徐立安等, 1999)。2000年1月调查5年生测定林的树高、胸径、冠幅; 2003年4月调查8年生测定林分的树高、胸径、冠幅、通直度, 并于胸高部位取木芯。
木材基本密度测定采用Smith(1954)的最大含水量法(李火根等, 1993); 管胞长度采用硝酸法离析, 经番红染色后在投影显微镜(×40)下随机测量50根, 取平均值; 管胞宽度在普通显微镜(×45)下随机测量30根管胞宽度, 取平均值。
1.2.2 统计分析方法试验中有关数据的方差统计分析均采用SAS(6.12版)(惠大丰等, 1996)完成。
方差分析模型为:Yij=μ+bi+fj+bfij+εij。式中:Yij为第i个家系第j个区组的观察值; μ为半同胞家系的平均效应; bi为第i个区组的效应; fj为第j个半同胞家系的效应; bfij为第i个半同胞家系和第j个区组的交互效应; εij为第i个家系第j个区组的观察值的随机误差。
表型变异系数(CV)和遗传变异系数(GCV)按公式: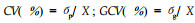
遗传力估算:
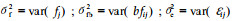
遗传增益估算: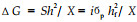
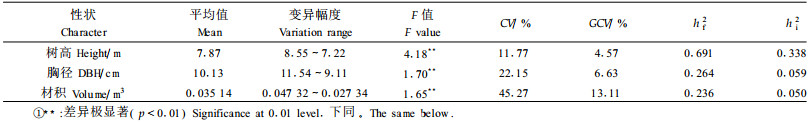
从表 1可以看出, 树高、胸径、材积性状在家系间差异极显著, 表明马尾松无性系种子园的半同胞家系子代林生长性状存在较丰富变异, 具有改良潜力。表型变异系数(CV)以材积为最大(45.27%), 其遗传变异系数(GCV)也最大(13.11%); 树高的CV和GCV最小, 分别为11.77%和4.57%。主要性状的CV和GCV都有以下的大小关系:材积>胸径>树高, 子代林5年生时的数据分析结果也与之一致。但各性状的遗传力大小关系则不同, 家系遗传力和单株遗传力的大小关系均为:树高>胸径>材积, 其中树高的家系遗传力为0.691, 属于强度遗传范围。
|
|
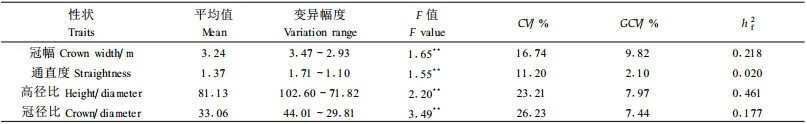
作为用材树种的良种应该满足材积生长量大、树干通直圆满、侧枝细小而平展、幼龄材基本密度高而且材性均匀等。在形质指标当中, 包含了通直度、圆满度、弯曲度、高径比、树皮指数、枝粗指数、自然整枝等多项指标, 考虑到对所有形质指标的调查有一定难度, 本次调查仅涉及冠幅、通直度、高径比(cm/cm)、冠径比(cm/cm)4个性状。
从表 2可见, 冠幅、通直度、高径比、冠径比在家系间差异均达极显著, 说明4个性状具有选择潜力。4个性状的CV也都达到10%以上的水平; GCV以冠幅的最大(9.82%), 通直度最小(2.1%); 遗传力以通直度相对较小, 仅为0.020, 属于弱遗传力水平, 仅相当以往报道(徐立安等, 1999)时的单株遗传力, 偏低。在高径比、冠径比两性状中, 高径比的家系遗传力比冠径比大很多, 进一步说明了树高比冠幅具有更高的遗传力。由于森林经营最终目的为木材产量, 因此在追求单株最大材积的同时, 还应注意单位面积的蓄积量, 这就要求考虑造林密度的问题, 而形质指标是影响造林密度的主要因素。所以在家系选择过程中, 要求材积生长量大, 窄冠, 大的高径比, 小的冠径比, 通直度良好者。
|
|
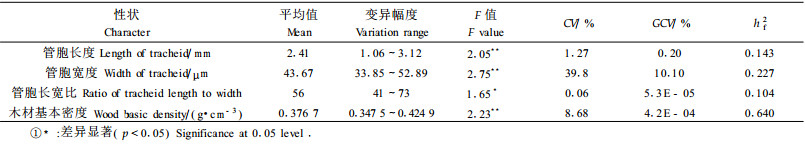
木材基本密度(g·cm-3)和管胞性状作为针叶树材性的重要指标, 在纸浆造纸工业中不仅影响制浆得率, 而且影响到纸张质量, 因此在针叶材改良中具有重要地位(施季森等, 1992)。表 3表明管胞长度(mm)和管胞宽度(μm)在半同胞家系间差异均为极显著, 管胞长宽比差异显著, 这些性状在家系内均为差异不显著, 说明马尾松半同胞家系管胞性状选择会有一定效果, 但管胞性状的遗传力较低。木材基本密度在半同胞家系间差异极显著, 半同胞家系间变异幅度也比较大(0.347 5~0.424 9), CV为8.68%, 而半同胞家系内的平均表型变异系数为7.5%, CV范围0.031~0.176, 家系内的木材基本密度差异不显著(F值为0.82, 概率为0.575 3);家系遗传力为0.640, 属于强度遗传, 与以往马尾松天然林优树子代的研究结果一致1), 也与Loo等(1984)研究火炬松(P. taeda)的结论相接近。表明木材基本密度的遗传力较大, 改良潜力较大。据此分析可以认为马尾松木材基本密度在不同家系间的选择将是有效的。
|
|
1) 邱龙广. 1991.马尾松天然林优树自由授粉子代木材基本密度遗传变异的研究.见:林木改良讨论会文集(杉木、马尾松等树种).北京:中国林学会遗传育种学会, 222-227
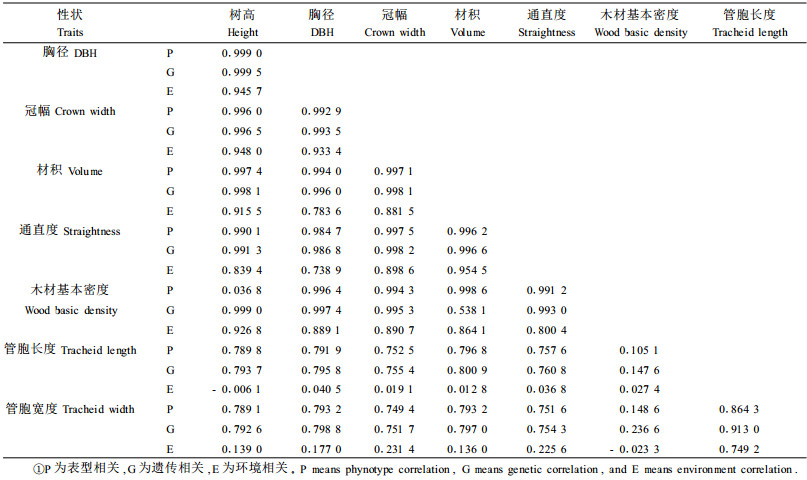
2.2 性状相关分析 2.2.1 相关分析生长量、形质和材质等作为林木常规育种的重要性状指标, 研究其相互间的相关性, 可以为间接选择提供一定的依据。从表 4可见, 8年生马尾松半同胞家系的树高、胸径、冠幅、材积之间的相关都达到极显著水平。通直度与木材基本密度、管胞长度、管胞宽度等材质性状的遗传相关达到极显著水平, 说明通过对树木干形(通直度)的选择会促进材质的改进, 即通过选择良好通直度的林木可间接改良材性。树高与木材基本密度的表型相关不显著; 树高、胸径、冠幅、材积与木材基本密度、管胞长度、管胞宽度等材质指标的遗传相关均达到极显著水平。木材基本密度与管胞长和宽两性状相关不显著。
|
|
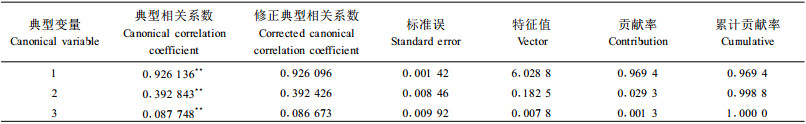
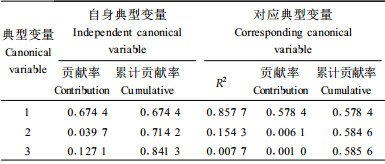
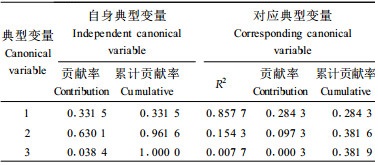
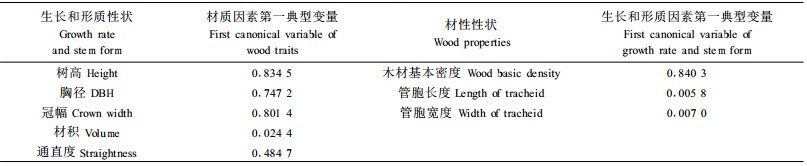
进一步剖析生长和形质性状与材性性状的相关性, 将树高、胸径、冠幅、材积、通直度5个性状指标作为第1组变量, 将材性指标中3个性状(木材基本密度、管胞长度、管胞宽度)作为第2组变量进行典型相关分析(表 5)。可见, 典型变量之间的第一典型相关系数为0.926 136, 达到了极显著, 且其相关信息占性状间总相关信息的96.94%。用它分析就可以完全代表二者之间的相互关系。分析生长和形质性状对材性性状(表 6)及材性性状对生长和形质性状的贡献率(表 7)发现, 生长和形质性状第一典型变量对材性性状变异的贡献率达到57.84%, 但材性性状第一典型变量却仅能解释生长和形质性状变量的28.43%。多元决定系数分析结果(表 8)表明, 材质因素第一典型变量当中只有木材基本密度对生长和形质性状有较好的预测作用, 决定系数达0.840 3, 而生长因素第一典型变量中除材积外, 其他均有一定的预测能力。这也补充说明上面相关分析中生长指标与材性指标相关密切情况:木材基本密度和树高、胸径、冠幅、材积的遗传相关显著。考虑到木材基本密度在纸浆造纸工业中的重要性, 在马尾松制浆造纸材改良过程中, 应注重生长量和木材基本密度两性状的改良。而本次分析表明马尾松树高生长在改良过程中, 有着尤为重要的地位, 因此初步认为在马尾松制浆造纸材改良中应适当顾及树高生长量。
|
|
|
|
|
|
|
|
比较不同家系树高和材积与对照的差异情况(表 9), 在5年生时, 以树高作为评价指标, 参试家系中有55.5%大于对照, 其中大于对照10%以上的家系仅占1.7%;以材积作为评价指标, 参试家系中有64.7%大于对照, 其中超过对照10%以上的家系占26.1%, 超过对照20%的占6.7%, 超过对照30%的仅占0.8%。在8年生时, 以树高作为评价指标, 参试家系中有37.8%大于对照, 无家系超过对照10%以上; 以材积作为评价指标, 参试家系中有55.5%大于对照, 其中超过对照10%以上的家系占22.7%, 超过对照20%的占4.2%, 超过对照30%的仅占0.8 %。可见, 以材积和树高生长量作为评选指标其选择结果显然存在一定差异, 这种差异还会随着林分年龄的增长而发生变化。因此在选择过程中应兼顾之。
|
|

若按短轮伐期经营, 不同选择强度下生长量性状的遗传增益估算结果见表 10, 可见5年生与8年生家系在相同选择强度下的遗传增益是比较接近的; 同时比较入选家系情况, 发现30%的入选率时有6个家系不同, 而20%入选率时仅1个家系不同, 10%时均相同。因此初步认为作为制浆造纸材短轮伐期经营时, 5年生时可作为优良家系选择的初选年龄。为了验证此结论的准确性, 进一步分析了5年生与8年生林分的树高、胸径与材积3个生长量性状的相关性, 发现3个性状的遗传相关系数分别达0.923 4、0.637 2和0.793 5, 相关程度均达极显著水平, 证实5年生可作为初选年龄。顾及种子园内无性系的开花结实情况(陈敬德, 2000), 按照10%的入选率, 有12个家系可入选。而比较树高和材积性状的入选结果, 发现659、8006、1、662、168、265和335共7个家系的树高和材积均在前12名之列, 15、509、14、658和11号家系虽树高生长在前12名, 但材积生长不如246、12、2、341和773号家系, 而后者的树高生长均排在前25名。综合考虑上述情形, 认为659、8006、1、662、168、265、335、246、12、2、341和773共12个家系属树高和材积生长优良者, 且这些家系的材质性状(包括密度、管胞长度和宽度)均排名于前43名, 因此可作为制浆造纸材短轮伐期的优良家系。
|
|

马尾松无性系种子园半同胞家系的生长量、形质和材性等性状在家系间存在极显著差异, 说明可以从参试家系中进一步选择得到生产力高的速生丰产优质的种质; 同时也表明根据表型选择得到的优树建立的无性系种子园其后代分化明显, 表型优良并不意味基因型一定优良。从表 9可见119个参试家系, 仍然有相当一部分家系生长量还不如对照。因此必须经过遗传测定, 才能选出优良基因型的种质。这一结论与马尾松的优树子代测定结果(全国马尾松种子园课题协作组, 1990; 周志春等, 1994; 徐立安等, 1997; 1999;郑仁华等, 1998; 龙光生等, 2002)一致。从表 9还可看出, 随着子代林分年龄的增长, 超过对照的参试家系数量似乎呈下降趋势, 这可能与本次测定设置的对照有关, 由于闽西地区马尾松属优良种源区(徐立安等, 1999), 而种子园的材料来自于福建全省的选优材料, 基因型与环境的互作问题可能是其中的一方面原因。考虑到种子园的种子要销往全省各地, 因此有必要建立多地点的测定林分, 揭示基因型与环境的互作问题, 来进一步综合评选已建无性系种子园内各无性系的遗传品质, 最终达到适地适种, 挖掘采种母树高效的遗传潜力。此外, 顾及我国种子园建设起步较晚, 为了加快高世代种子园营建的进程, 目前对已建种子园大多采取边测定边利用的原则。在此背景下, 种子园既是生产群体, 又起育种群体的作用。由此, 利用种子园材料可进一步研究群体的交配系统、基因流动、遗传多样性等重要理论问题, 并能为营建高世代种子园提供指导。而利用种子园的子代测定获得的遗传参数, 不仅可为解决种子园的理论问题提供帮助, 而且可有效评价园内母树的遗传背景, 从而可从中选择一般配合力高的材料, 进一步开展人工交配设计做全同胞测定, 为高世代育种服务。综上可见, 对马尾松种子园的无性系进行子代测定是极为必要的, 其现实和远景意义将不言而喻。
分别估算了8年生马尾松种子园半同胞子代树高、胸径、材积的家系和单株遗传力, 通直度、高径比、冠径比的家系遗传力。单株遗传力以树高为最高(0.338);家系遗传力的大小关系为:树高(0.691)>高径比(0.461)>胸径(0.264)>材积(0.236)>冠幅(0.218)>冠径比(0.177)>通直度(0.020), 即树高生长具有较强的遗传力, 而胸径和材积等受较弱遗传力控制。此结论与周志春等(2000)对马尾松优树13年生子代测定林的估算结果:树高、胸径均受中等至强度遗传力控制, 略有出入。
性状相关分析表明:8年生马尾松半同胞家系在生长量、形质和材性等主要性状之间的相关, 均达到了极显著水平; 通直度与树高等性状相关也达到极显著水平, 表明在改良树高、胸径、材积等性状的同时, 干形(通直度)也能得到一定的间接改良; 木材基本密度与管胞性状在半同胞家系间的相关不显著; 管胞长度与管胞宽度相关在家系间都达到了显著水平。本次试验的材积与材性方面达到显著正遗传相关, 与管兰华1)以杨树为材料的研究结果相一致; 木材基本密度与材积呈极显著正相关, 与范林元2)的马尾松实生种子园材料和徐立安等(1997)的马尾松种源子代材料研究结论不同。本次测定发现生长和形质性状与材质相关密切, 说明生长性状的改良有利于材质的改善。考虑到木材材质性状和生长性状的本身变异较大, 因此可以直接对那些木材基本密度较高、管胞长度与管胞宽度均佳、而生长又迅速的家系或个体进行选择, 这与以往马尾松种源水平的研究结果(王章荣, 1998)有一定差异。Zobel等(1989)指出树木生长速度与木材基本密度、纤维长度之间的相关性, 因树种、树龄和地理位置不同而不同, 有的明显相关, 有的不相关, 说明生长性状与材性指标的相关分析, 与参试的试验材料有关。因此在作间接选择时应注意这一点。
1) 管兰华. 2001.杨树新无性系生长材性联合遗传改良及其早期选择研究.南京林业大学, 硕士学位论文
2) 范林元. 2001.马尾松实生种子园亲本多性状育种值评定及其去劣疏伐研究.南京林业大学, 硕士学位论文
典型相关分析发现, 材质因素第1组变量中只有木材基本密度对生长性状有较好的预测作用, 决定系数达0.840 3, 而生长因素第1组变量中除材积外, 其他均有一定的预测能力。进一步可看出, 生长性状与木材基本密度相关显著, 这有利于制浆造纸材的选育。众所周知, 制浆造纸材选育的目标是追求高得浆率, 生长量和木材基本密度能直接影响到单位造林面积上的得浆总量, 若生长性状与木材基本密度相关显著, 在制浆造纸材选育过程中, 只要直接考虑易于观测到的生长性状, 而木材基本密度的指标可间接获得。
根据性状遗传力和性状间的相关性分析结果, 顾及性状调查和分析的直观性, 认为在马尾松制浆造纸材优良家系选择时, 可适当注重树高生长优势者。
在10%入选率下, 5年生林分的选择结果与8年生的选择结果一致, 且5年生与8年生林分的树高、胸径与材积3个生长量性状的遗传相关均达极显著。因此认为在马尾松制浆造纸材短轮伐期经营过程中, 5年生时可作为优良制浆造纸材家系选择的初选年龄。与王章荣等(1999)以短轮伐期为经营模式的优良种源选择结论基本一致。而从子代测定林总体长势的实地考察来看, 也确实发现有些家系一直处于中庸甚至劣势, 有必要尽早对之进行淘汰。当然, 若以家系测定结果来指导种子园的疏伐, 在尽可能伐除中庸者之际, 还得综合考虑种子园无性系的开花结实性状, 毕竟种子园的最终经营目标是生产经过遗传改良的种子。因此在种子园疏伐过程中, 应同时兼顾母树遗传品质和雌雄花量问题, 使种子园真正达到高产优质。
据制浆造纸材短轮伐期的思路, 以树高和材积生长量作为选择指标, 按照10%的入选率, 兼顾种子园无性系的开花结实情况, 筛选出优良家系12个, 它们的材质排名于前43名, 可作为制浆造纸材优良家系。
陈敬德. 2000. 马尾松无性系种子园抚育疏伐效果分析. 林业科技开发, 14(4): 19-20. |
惠大丰, 姜长鉴. 1996. 统计分析系统SAS软件实用教程. 北京: 北京航空航天大学出版社.
|
季孔庶. 2001. 马尾松人工林培育技术. 北京: 中国农业出版社, 14.
|
李火根, 王章荣, 陈天华. 1993.马尾松不同种源林分内个体间木材性状的变异.见: 涂忠虞, 沈熙环主编.中国林木遗传育种进展.北京: 科学技术文献出版社, 27-32
|
龙光生, 李午平, 葛宜和, 等. 2002. 马尾松半同胞优良家系选择研究. 中南林学院学报, 22(1): 17-22. DOI:10.3969/j.issn.1673-923X.2002.01.004 |
全国马尾松种子园课题协作组. 1990. 马尾松种子园建立技术论文集. 北京: 学术书刊出版社, 155-203, 275-313.
|
施季森, 叶志宏, 翁玉榛, 等. 1992.杉木生长与材性联合遗传改良研究.见: 沈熙环主编.种子园技术.北京: 北京科学技术出版社, 78-92
|
王章荣, 陈天华, 周志春, 等. 1999. 马尾松制浆造纸性能的群体变异及适应性试验. 林产化学与工业, 19(1): 64-68. DOI:10.3321/j.issn:0253-2417.1999.01.014 |
王章荣. 1998. 马尾松制浆造纸材定向选育. 林业科技开发, 12(2): 6-8. |
徐立安, 陈天华, 王章荣, 等. 1997. 马尾松种源子代材性变异与制浆造纸材优良种源选择. 南京林业大学学报, 21(2): 1-6. |
徐立安, 王章荣, 曹汉洋. 1999. 福建省马尾松优树自由授粉子代变异及分析. 福建林学院学报, 19(2): 114-117. DOI:10.3969/j.issn.1001-389X.1999.02.005 |
郑仁华, 蔡天贵, 陈国金, 等. 1998. 马尾松优树子代测定及速生优良家系选择的研究. 福建林业科技, 25(3): 11-16. DOI:10.3969/j.issn.1002-7351.1998.03.003 |
周志春, 金国庆, 周世水. 1994. 马尾松自由授粉家系生长和材质的遗传分析及联合选择. 林业科学研究, 7(3): 253-257. DOI:10.3321/j.issn:1001-1498.1994.03.001 |
周志春, 李光荣, 黄光霖, 等. 2000. 马尾松木材化学组分的遗传控制及对木材育种的意义. 林业科学, 36(2): 110-115. DOI:10.3321/j.issn:1001-7488.2000.02.018 |
Loo J A, Tauer C G, van Buijtenen J P. 1984. Juvenile-mature relationships and heritability estimates of several traits in loblolly pine (Pinus taeda). Can J For Res, 14: 822-825. DOI:10.1139/x84-145 |
Zobel B J, van Buijtenen J P. 1989. Wood variation:its causes and control. Berlin: Springer-Verlag, 249-290.
|