 2005, Vol. 41
2005, Vol. 41文章信息
- 李昌晓, 钟章成.
- Li Changxiao, Zhong Zhangcheng.
- 模拟三峡库区消落带土壤水分变化条件下落羽杉与池杉幼苗的光合特性比较
- Comparative Studies on Photosynthetic Characteristics of Taxodium distichum and Taxodium ascendens Seedlings under Simulated Soil Water Change in the Hydro-Fluctuation Belt of Three Gorges Reservoir Area
- 林业科学, 2005, 41(6): 28-34.
- Scientia Silvae Sinicae, 2005, 41(6): 28-34.
-
文章历史
- 收稿日期:2005-04-18
-
作者相关文章



三峡库区消落带系水陆系统之间的过渡性生态系统(黄京鸿, 1994; 刁承泰等, 1999; 张洪江等, 2000), 对于三峡库区库岸安全以及三峡水库的正常运行具有重要生态保障作用。根据当地实际情况, 尽快恢复好消落带植被, 切实建设好库岸防护林体系, 无疑是三峡库区消落带能够发挥正常生态保障功能的关键。三峡库区消落带因水位周期性常年变化, 使得土壤含水量呈现出从干旱状态到全水淹状态的一系列梯度性变化。消落带土壤含水量的这种梯度性变化势必影响到植物的生长发育及其生理生态学特性, 尤其是光合特性, 从而对消落带适生植物种类提出了更高的要求。因此选出适于在三峡库区消落带这种水分环境复杂多变条件下生长的最佳适生树种不仅尤为重要, 而且意义重大。目前已有大量土壤水分变化条件下林木生理生态特性的报道(何维明等, 2000; 陈德祥等, 2003; 刘伟玲等, 2003; 马成仓等, 2004; 史胜青等, 2004; Anderson et al., 1999; Simone et al., 2003), 但是缺乏有关三峡库区消落带土壤水分变化条件下林木光合生理生态特性的报道。本研究的目的是通过模拟三峡库区消落带土壤水分变化特征, 比较研究适生树种的光合生理生态学特性, 为选择出最适于三峡库区消落带植被恢复建设的造林绿化树种提供依据。
1 材料与方法 1.1 研究树种和地点对生物学特性相近的落羽杉(Taxodium distichum)和池杉(T. ascendens)当年实生幼苗的光合生理生态特性进行比较研究。2004年6月中旬将落羽杉和池杉生长基本一致的各120株幼苗带土盆栽(土壤为紫色土), 每盆1株。盆中央内径为13 cm, 盆内土层厚度12 cm。将所有盆栽试验用苗置于西南师范大学生态试验园地中(海拔249 m)进行相同土壤基质、光照和水分管理, 于7月25日搭建透明塑料遮雨棚, 开展试验。
1.2 试验设计将2种试验用苗各随机分成4组(对照组CK、轻度干旱组T1、水分饱和组T2和水淹组T3), 每组30盆。对照组CK即为常规生长组, 土壤含水量(采用称重法测定)为田间持水量的60%~63%, 盆栽幼苗在晴天无萎蔫现象(郭连生等, 1994; 胡新生等, 1998; 胡哲森等, 2000)。轻度干旱组T1即为轻度水分胁迫, 土壤含水量为田间持水量的47%~50%, 植株嫩叶在晴天13:00左右出现萎蔫, 17:00左右恢复正常(郭连生等, 1994; 胡新生等, 1998; 胡哲森等, 2000)。水分饱和组T2即为表面一直处于潮湿状态的水饱和土壤。水淹组T3在本试验中为苗木根部土壤全部淹没, 淹水超过土壤表面1 cm。水淹处理时, 将苗盆放入直径为68 cm、高22 cm的大型塑料盆内, 然后向盆内注水, 直到盆内水面超过土壤表面1 cm为止(Bragina et al., 2001)。
从试验处理之日算起, 每间隔5 d为一个处理期, 对各项生理生化指标进行连续测定, 即分别在7月31日、8月6日、8月12日、8月18日和8月24日开展5次测定, 每个处理每次测定5个重复, 最后取5次测定的平均值作为比较结果。8月25日结束试验。
1.3 叶片交换气体参数的测定在预备试验的基础上, 选取2种苗木从上往下数的第3至第4片叶在饱和光强下完成光诱导后, 使用美国CID公司生产的CI-310 POS便携式光合系统直接测定叶片气体交换系数。所有测定均在9:00—11:00内25 ℃的室温环境下完成。每次在400 μmol·L-1 CO2和1 000 μmol photons·m-2 s-1光强下测定叶片的净光合速率。测定参数包括净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、气温(ta)、叶温(tleaf)、空气相对湿度(RH)、胞间CO2浓度(ci)。水分利用效率(WUE)=Pn/Tr (Nijs et al., 1997)、表观光能利用效率(LUEapp )=Pn/PAR (Long et al., 1993)、表观CO2利用效率(CUEapp)=Pn/ci(何维明等, 2000)。
1.4 光合色素含量的测定采用浸提法(邹琦, 1995)用岛津5220分光光度计测定叶绿素(Chls)、类胡萝卜素(Car)含量。
1.5 统计分析根据测定生理指标, 将水分处理作为独立因素, 用一元方差分析(One-Way ANOVA)揭示落羽杉和池杉在水分变化条件下的光合生理生态学特性(GLM程序, SPSS 10.0版), 并用Duncan检验法进行多重比较, 检验2树种幼苗每个生理指标在处理间(α=0.05)的差异显著性(杜荣骞, 2003)。
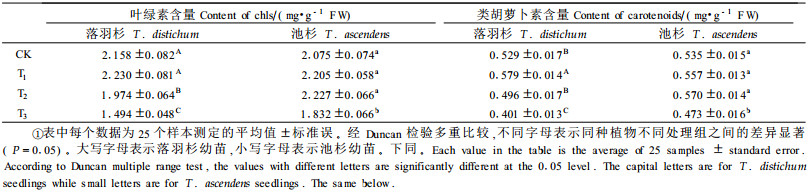
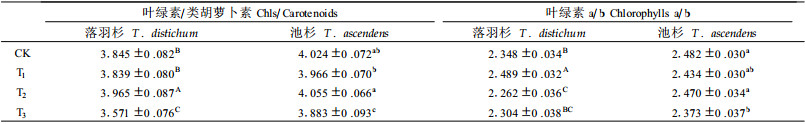
2 结果 2.1 光合色素的变化方差分析结果(表 1、2、3)显示, 不同水分处理均能显著影响落羽杉和池杉幼苗的光合色素含量。经Duncan检验多重比较, 发现落羽杉和池杉幼苗水淹处理组T3的光合色素含量平均值在各自处理组均处于最低, 与对照组CK以及轻度干旱组T1和水分饱和组T2均有极显著差异。但是, 落羽杉幼苗光合色素含量在轻度干旱以及正常生长条件下与相同处理条件的池杉幼苗光合色素含量并无显著差异性; 相反, 在饱和水以及水淹条件下落羽杉幼苗光合色素含量始终小于池杉, 且二者光合色素含量差异达到极显著。在整个试验期, 落羽杉幼苗叶绿素a与b的比值介于2.043~2.691之间波动, 叶绿素与类胡萝卜素的比值则介于3.079~4.514之间波动; 池杉幼苗叶绿素a与b比值介于2.124~2.644之间, 叶绿素与类胡萝卜素之比介于3.331~4.387之间。2树种的变化范围基本一致。
|
|
|
|

|
|
通过方差分析得知, 不同水分变化能极其显著地影响落羽杉和池杉幼苗净光合速率Pn、蒸腾速率Tr和气孔导度Gs(图 1)。经Duncan检验多重比较, 发现落羽杉幼苗Pn平均值在T2和T3组以及与CK组之间并无显著差异; T1组却极显著地低于其他3组, 其净光合速率比正常下降24.949%。池杉幼苗Pn平均值在T3和T2组之间无显著差异, 分别高出CK组3.238%和9.421%;与落羽杉幼苗一样, 池杉幼苗T1组Pn平均值也最低, 与另外3组均存在极显著差异, 低于CK组23.062%。落羽杉幼苗T2组Pn平均值显著低于池杉幼苗T2组的Pn平均值, 降低幅度达12.466%(P=0.014<0.05), 与CK组二者Pn平均值无显著差异形成鲜明对比(P=0.607>0.05)。

|
图 1 落羽杉和池杉幼苗在不同水分条件下净光合速率、蒸腾速率和气孔导度的变化(±标准误) Fig. 1 The change of Pn, Tr and Gs of T. distichumand T. ascendens seedlings under different water treatment (±SE) 根据不同树种每个处理测定的25个样本平均值作图, 误差棒为平均值的标准误。经Duncan检验多重比较, 不同字母表示同种植物不同处理组之间的差异显著(P=0.05)。大写字母表示落羽杉幼苗, 小写字母表示池杉幼苗。下同。 Each column stands for the mean of 25 samples, and the error bar represents standard error of the mean. According to Duncan multiple range test, the values with different letters are significantly different at the 0.05 level. The capital letters are for T. distichum seedlings while small letters are for T. ascendens seedlings. The same below. |
落羽杉幼苗的蒸腾速率Tr和气孔导度Gs平均值在各处理组之间均随着土壤水分含量的增加而显著递增, 即T1组最小, T3组最大。池杉幼苗T2与T3组的Tr和Gs平均值虽然也均大于CK组, 且与CK组有极显著差异, 但4组中以T2组平均值最大; T1组平均值尽管小于CK组, 但与CK组并无显著差异。两树种幼苗Tr和Gs平均值在单个处理组内分别比较, 发现在T3组落羽杉幼苗的气孔导度要显著大于池杉幼苗(P=0.024<0.05), 而在CK组不论是Tr还是Gs平均值均无显著差异(Tr : P=0.884>0.05;Gs: P=0.612>0.05)。
2.3 资源利用效率的变化方差分析结果可知, 不同水分处理对落羽杉和池杉幼苗资源利用效率的影响均极其显著(图 2)。在整个试验期, 落羽杉幼苗CK组的WUE平均值最高, 另3组则随着土壤水分含量增加而降低。池杉幼苗T2和T3组在整个试验期的WUE平均值并无显著差异, 但显著低于T1组和CK组, 并且也以CK组的WUE平均值最高(图 2)。同时2树种幼苗在CK组的WUE 平均值并无显著差异(P=0.324>0.05)。

|
图 2 落羽杉和池杉幼苗水分利用效率、表观光能利用效率和表观CO2利用效率的变化(±标准误) Fig. 2 The WUE, LUEapp and CUEapp of T. distichum and T. ascendens seedlings under different water treatment (±SE) |
落羽杉幼苗T1组的LUEapp和CUEapp平均值与CK组以及T2、T3组均有显著差异, 但T2、T3、CK组相互之间并无显著差异。池杉幼苗LUEapp和CUEapp的平均值在T2和T3组之间无显著差异, 但均高于CK组和T1组。经Duncan检验多重比较, 发现落羽杉幼苗LUEapp和CUEapp在T2组平均值均显著小于相同处理组池杉幼苗平均值(PLUE=0.015<0.05, PCUE= 0.005<0.01), 而在CK组2树种幼苗LUEapp和CUEapp的平均值均差异不显著(PLUE=0.638>0.05, PCUE=0.780>0.05)。
2.4 相关性分析相关性分析结果(表 4)显示, 落羽杉幼苗Pn与Tr、Gs、WUE、LUEapp以及CUEapp均达到极显著正相关, 而与叶绿素(Chls)、类胡萝卜素(Car)含量以及二者的比值(Chls/Car)、大气相对湿度和胞间CO2浓度均无显著相关关系, 但与叶绿素a/b的比值表现出极显著负相关。
|
|
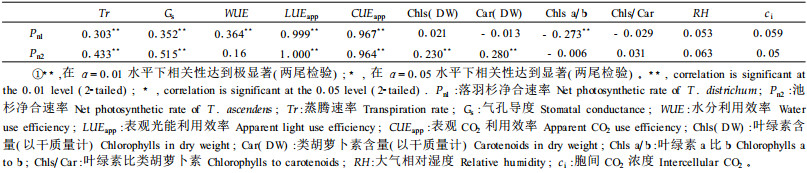
池杉幼苗Pn与Tr、Gs、LUEapp、CUEapp、叶绿素和类胡萝卜素含量(以干质量计)均达极显著或显著正相关, 而与WUE、叶绿素含量(以鲜质量计)、类胡萝卜素含量(以鲜质量计)、叶绿素与类胡萝卜素比值、叶绿素a与b比值、大气相对湿度、胞间CO2浓度并未表现出直接显著相关性。
3 讨论三峡库区消落带周期性水位变化在对消落带现有植物带来一系列生理生态学适应性改变的同时, 对未来适生植物的耐水湿性能也提出了更高要求。落羽杉和池杉均属裸子植物杉科(Taxodiaceae)落羽杉属(汪企明等, 1995; 1998), 原产于北美东南部沼泽地区, 通常具有膝状呼吸根(陶仁中等, 1998; 郑松发等, 2004), 耐水湿性能均很强, 现在我国长江流域多有引种(汪贵斌等, 2002)。
本试验研究发现落羽杉幼苗在过多水分条件下(如T2、T3组)光合色素合成数量要远低于池杉幼苗, 从一个侧面证实了落羽杉幼苗耐水淹与渍水能力可能小于池杉幼苗。一般而言, 正常叶子叶绿素与类胡萝卜素比值、叶绿素a与b比值均约为3:1(潘瑞炽等, 2004)。落羽杉和池杉幼苗叶绿素与类胡萝卜素比值大于3:1既可起到提高叶绿素在光合色素中相对含量比例进而增加光合能力的作用, 又可同时起到确保有足够反应中心色素的作用。2种植物叶绿素a与b的比值始终小于3:1, 可起到保证有充足的聚光色素参与光能合成的作用, 使叶绿素a与b的分配比例显得更加合理高效(史胜青等, 2004; Scholes et al., 1997; Ronzhina et al., 2004)。
在整个试验期, 落羽杉幼苗Pn平均值在土壤饱和水与水淹处理条件下与对照组基本一致, 其光合能力并未受到任何影响(图 1), 可以证实落羽杉树种具有较强的耐水湿生理生态学特性, 这与前人的研究结论完全一致(汪贵斌等, 2004; Conner et al., 1997; Pezeshki et al., 1998; Middleton, 2000; Eclan et al., 2002)。然而, 与落羽杉幼苗有所不同, 池杉幼苗Pn平均值在土壤饱和水与水淹处理条件下均高于对照组, 其光合能力得到明显加强, 由此可以推断池杉幼苗比落羽杉幼苗可能具有更强的耐水湿能力(李兆玉, 1993)。特别是在饱和水条件下, 池杉幼苗的净光合速率平均值最高, 显示出土壤水分饱和是池杉幼苗的最适生长条件(何维明等, 2000)。在干旱水分胁迫环境条件下, 落羽杉与池杉幼苗净光合速率均显著偏低, 说明土壤水分供应不足对二者光合作用净产物积累均会产生明显的负面影响(喻方圆等, 2004)。
轻度干旱的水分胁迫条件并未降低池杉幼苗的蒸腾速率和气孔导度, 这意味着在轻度干旱条件下池杉幼苗仍然保持着蒸腾速率和气孔导度的正常水平。由此可以证明池杉幼苗不仅具有耐水淹和渍水的能力, 还具有一定程度的耐旱性。由于落羽杉幼苗并未表征出相应的耐旱特性, 因而有理由推断池杉幼苗具有比落羽杉幼苗更为宽泛的水分胁迫适应范围(吴祖映等, 1996)。池杉幼苗在土壤水分饱和条件下的Tr和Gs平均值显著大于另外3组, 进一步说明土壤水分饱和是池杉幼苗的最适生长条件, 这与落羽杉和池杉的生物学特性应该有很大关系(汪企明等, 1995)。
试验结果表明, 落羽杉水淹处理组水分利用效率低于水分饱和处理组25.482%, 与池杉水淹处理组同水分饱和处理组无显著差异形成鲜明对比, 由此可以推知池杉幼苗在水淹处理条件下水分利用效率降低的敏感表征度并不如落羽杉表征明显(殷云龙等, 1997)。落羽杉幼苗渍水和水淹环境下具有与对照组基本一致的表观光能利用效率和表观CO2利用效率, 与池杉幼苗在相同条件下具有高于对照组的表观光能和表观CO2利用效率形成明显对比, 更进一步说明水淹和渍水的环境条件更加有利于池杉幼苗的生长发育(郑松发等, 2004)。轻度干旱条件下2树种表观光能和表观CO2利用效率显著低于其他各组说明水分亏缺均不利于它们的生长发育(曾慧卿等, 1999)。
由于轻度干旱组和水饱和、水淹2个处理组处于正常生长条件所需水分的2个不同发展方向, 并非正常生长条件之上或之下的单向梯度性水分设定, 因而使得总体相关性分析结果可能出现不能完全代表单个处理组内各参数之间的相关情况。故对相关性的分析还需要进一步考虑各组的具体情况。如池杉幼苗净光合速率Pn与水分利用效率WUE虽然并未从整体上表现出显著相关性, 但方差分析检验发现池杉幼苗CK组的Pn与WUE呈现出极显著正相关(P=0.000<0.001, R=0.778***), 且Pn与WUE遵循二次多项式Pn=-0.007 2(WUE)2+0.334 7(WUE)+1.518 5。
从本试验研究结果来看, 落羽杉和池杉树种均可考虑列为三峡库区消落带的防护林体系建设树种。在栽植营造以及管护时, 必须充分考虑到落羽杉和池杉树种的耐水湿性特点, 将它们置于土壤水分含量充足的环境之中, 尤其是将池杉置于土壤饱和水的环境里生长效果最佳。但是落羽杉树种不宜置于干旱环境之中; 相反池杉树种对适应水分逆境条件具有较高的可塑性, 不仅表现出耐水湿的特点, 还表征出一定程度的耐旱性, 可以适当置于轻度干旱环境条件下, 但应注意浇水抗旱, 使池杉保持正常的净光合速率。
陈德祥, 李意德, 骆土寿, 等. 2003. 海南岛尖峰岭热带山地雨林下层乔木中华厚壳桂光合生理生态特性的研究. 林业科学研究, 16(5): 540-547. DOI:10.3321/j.issn:1001-1498.2003.05.004 |
刁承泰, 黄京鸿. 1999. 三峡水库水位涨落带土地资源的研究. 长江流域资源与环境, 8(1): 75-80. |
杜荣骞. 2003. 生物统计学. 第二版. 北京: 高等教育出版社, 104-116.
|
郭连生, 田有亮. 1994. 四种针叶幼树的光合生理特性与大气湿度关系的研究. 生态学报, 14(2): 136-141. DOI:10.3321/j.issn:1000-0933.1994.02.001 |
何维明, 马风云. 2000. 水分梯度对沙地柏幼苗荧光特征和气体交换的影响. 植物生态学报, 24(5): 630-634. DOI:10.3321/j.issn:1005-264X.2000.05.021 |
胡新生, 王世绩. 1998. 树木水分胁迫生理与耐旱性研究进展及展望. 林业科学, 34(2): 77-88. DOI:10.3321/j.issn:1001-7488.1998.02.011 |
黄京鸿. 1994. 三峡水库水位涨落带的土地资源及其开发利用. 西南师范大学学报(自然科学版), 19(5): 528-533. |
胡哲森, 许长钦, 傅瑞树. 2000. 锥栗幼苗对水分胁迫的生理响应及6-BA的作用. 福建林学院学报, 20(3): 1-4. |
李兆玉. 1993. 里下河地区不同树种耐水性的调查. 江苏林业科技, 20(1): 28-29. |
刘伟玲, 谢双喜, 喻理飞. 2003. 几种喀斯特森林树种幼苗对水分胁迫的生理响应. 贵州科学, 21(3): 51-55. DOI:10.3969/j.issn.1003-6563.2003.03.013 |
马成仓, 高玉葆, 王金龙, 等. 2004. 内蒙古高原甘蒙锦鸡儿光合作用和水分代谢的生态适应性研究. 植物生态学报, 28(3): 305-311. DOI:10.3321/j.issn:1005-264X.2004.03.004 |
潘瑞炽, 王小菁, 李娘辉. 2004. 植物生理学. 第五版. 北京: 高等教育出版社, 66-68.
|
史胜青, 袁玉欣, 杨敏生, 等. 2004. 水分胁迫对4种苗木叶绿素荧光的光化学淬灭和非光化学淬灭的影响. 林业科学, 40(1): 168-173. DOI:10.3321/j.issn:1001-7488.2004.01.028 |
陶仁中, 江泽慧, 费本华, 等. 1998. 间歇性淹水对池杉木材物理力学性质影响的研究. 林业科学, 34(3): 110-114. DOI:10.3321/j.issn:1001-7488.1998.03.016 |
汪贵斌, 曹福亮. 2002. 落羽杉抗性研究综述. 南京林业大学学报(自然科学版), 26(6): 78-82. DOI:10.3969/j.issn.1000-2006.2002.06.020 |
汪贵斌, 曹福亮. 2004. 土壤盐分及水分含量对落羽杉光合特性的影响. 南京林业大学学报(自然科学版), 28(3): 14-18. DOI:10.3969/j.issn.1000-2006.2004.03.004 |
汪企明, 江泽平, 吕祥生, 等. 1995. 落羽杉属种源研究:树种生物学特性. 江苏林业科技, 22(2): 14-18. |
汪企明, 王伟, 蒋志新, 等. 1998. 落羽杉属种源研究:生长和生物量变异. 江苏林业科技, 25(1): 1-6. |
吴祖映, 储家淼, 唐明荣, 等. 1996. 土壤水分状况对池杉形态结构及生长状况的影响. 浙江林学院学报, 13(3): 364-366. |
殷云龙, 陈永辉. 1997. 中山杉与池杉、落羽杉和水杉对比造林的调查和评价. 植物资源与环境, 6(3): 23-28. |
喻方圆, 徐锡增, Robert D Guy. 2004. 水分和热胁迫处理对4种针叶树苗木气体交换和水分利用效率的影响. 林业科学, 40(2): 38-44. DOI:10.3321/j.issn:1001-7488.2004.02.007 |
曾慧卿, 薛建辉. 1999. 江苏池杉人工林培育研究综述. 江苏林业科技, 26(3): 52-55. |
张洪江, 高中琪, 解明曙, 等. 2000. 三峡库区多功能防护林体系构成与布局的思考. 长江流域资源与环境, 9(4): 479-486. DOI:10.3969/j.issn.1004-8227.2000.04.013 |
郑松发, 陈玉军, 陈文沛, 等. 2004. 深水裸滩红树植物与落羽杉生长适应性研究. 林业科学研究, 17(5): 654-659. DOI:10.3321/j.issn:1001-1498.2004.05.017 |
邹琦. 1995. 植物生理学与生物化学试验指导. 北京: 中国农业出版社, 36-39.
|
Anderson P H, Pezeshki S R. 1999. The effects of intermittent flooding on seedlings of three forest species. Photosynthetica, 37(4): 543-552. |
Bragina T V, Martinovich L I, Rodionova N A, et al. 2001. Ethylene-induced activation of xylanase in adventitious roots of maize as a response to the stress effect of root submersion. Applied Biochemistry and Microbiology, 37(6): 618-621. DOI:10.1023/A:1012363319758 |
Conner W H, Mcleod K W, McCarron J K. 1997. Flooding and salinity effects on growth and survival of four common forested wetland species. Wetlands Ecology and Management, 5: 99-109. DOI:10.1023/A:1008251127131 |
Eclan J M, Pezeshki S R. 2002. Effects of flooding on susceptibility of Taxodium distichum L. seedlings to drought. Photosynthetica, 40(2): 177-182. DOI:10.1023/A:1021381204684 |
Long S P, Baker N R, Raines C A. 1993. Analyzing the responses of photosynthetic CO2 assimilation to long-term elevation of atmospheric CO2 concentration. Vegetation, 104/105: 33-45. DOI:10.1007/BF00048143 |
Middleton B. 2000. Hydrochory, seed banks, and regeneration dynamics along the landscape boundaries of a forested wetland. Plant Ecology, 146: 169-184. |
Nijs I, Ferris R, Blum H. 1997. Stomatal regulation in a changing climate: A field study using free air temperature increase (FATI) and free air CO2 enrichment. Plant, Cell and Environment, 20: 1041-1050. DOI:10.1111/j.1365-3040.1997.tb00680.x |
Pezeshki S R, Santos M I. 1998. Relationships among rhizosphere oxygen deficiency, root restriction, photosynthesis, and growth in baldcypress (Taxodium distichum L.) seedlings. Photosynthetica, 35(3): 381-390. DOI:10.1023/A:1006912318352 |
Ronzhina D A, Nekrasova G F, P'yankov V I. 2004. Comparative characterization of the pigment complex in emergent, floating, and submerged leaves of hydrophytes. Russian Journal of Plant Physiology, 51(1): 21-27. |
Scholes J D, Press M C, Zipperlen S W. 1997. Differences in light energy utilization and dissipation between dipterocarp rainforest tree seedlings. Oecologia, 109: 41-48. DOI:10.1007/s004420050056 |
Simone O D, Junk W J, Schmidt W. 2003. Central Amazon floodplain forests: Root adaptations to prolonged flooding. Russian Journal of Plant Physiology, 50(6): 848-855. DOI:10.1023/B:RUPP.0000003285.70058.4c |