 2005, Vol. 41
2005, Vol. 41文章信息
- 韩照祥, 山仑.
- Han Zhaoxiang, Shan Lun.
- 栓皮栎种群变异与适应对策研究
- Variation and Adaptive Countermeasures of Quercus variabilis Population in Shaanxi Province
- 林业科学, 2005, 41(6): 16-22.
- Scientia Silvae Sinicae, 2005, 41(6): 16-22.
-
文章历史
- 收稿日期:2003-09-01
-
作者相关文章


2. 淮海工学院化学工程系 连云港 222005;
3. 中国科学院 水利部水土保持研究所 杨凌 712100
2. College of Engineering of Huai Hai Lianyungang 222005;
3. Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resource Yangling 712100
栓皮栎(Quercus variabilis)是壳斗科(Fagaceae)栎属(Quercus)的落叶乔木, 喜光, 耐干旱瘠薄, 是我国暖温带落叶阔叶林和亚热带常绿落叶阔叶混交林的重要建群种, 也是一种经济价值很高的树种, 在发展经济、维护生态平衡等方面发挥着重要作用(中国树木志编委会, 1976)。由于栓皮栎受人为破坏的现象十分严重, 分布面积日益减少, 其种群质量和数量不断下降, 种群的稳定性受到严重破坏, 大多数地区栓皮栎种群有逐渐转移退缩的趋势。
栓皮栎具有很强的萌生能力, 萌生作为有性生殖的有益补充, 大大提高了栓皮栎种群抵御外界干扰和适应的能力(吴明作, 1998)。通过对栓皮栎实生苗及萌生苗的径级大小、数量、种群性状分化和内部基因层次等的测定和研究, 探讨栓皮栎种群沿生境梯度从分布核心区到分布边缘区的变异适应和进化规律, 为进一步研究栓皮栎种群对外界环境干扰的适应能力和种群生产力提供理论依据。
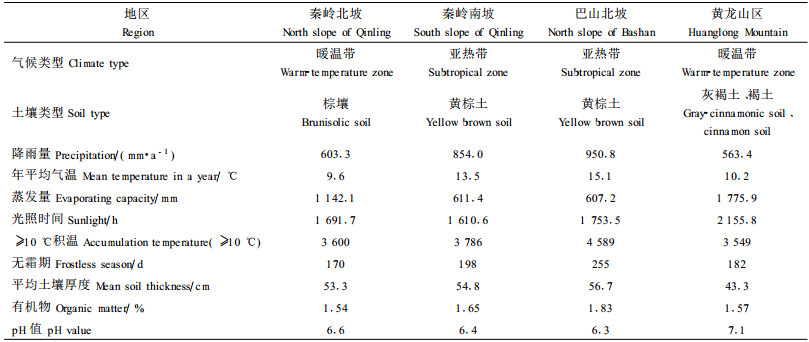
1 研究区自然概况和研究方法 1.1 研究区自然概况根据栓皮栎种群在陕西分布的特点, 将研究地设在栓皮栎种群分布中心的秦岭北坡(A样地)、秦岭南坡(B样地), 过渡地带的巴山北坡(C样地)和分布边缘地带的黄龙山区(D样地), 4个地区在气候类型、土壤和植被等方面存在很大差异(表 1)。
|
|
采用样方法进行野外调查, 在A、B、C、D 4样地分别选择生长发育中等、具有代表性的栓皮栎群落地段, 分别设置20 m×20 m的样方20个, 共设样方80个。对样地内的全部栓皮栎调查, 包括种类、高度、胸径、基株数、茎干数、冠幅及样地中种群个体数目等指标。
1.2.2 生态型性状分化参数的测定在生长季节分别在A、B、C、D 4样地中选择生长发育中等, 能代表各个样地生长状况的胸径为11、14、18、22、25 cm的植株各2株, 样本间距5 m以上, 测定每株的高度, 测量第1主枝长、枝间距、主枝倾角, 第2级、第3级的小枝长、倾角、叶倾角、分枝率等参数。叶倾角的测量采用实测和Warren-Wilson公式计算:α=arctgΠf13/2· f52, (f13、f52分别为样方倾角13°和52°的点样方测得的叶片接触频率), 叶方位角采用叶片上表面和法线取向, 用指南针(精度为0.1°)来测量(袁嘉祖等, 1991), 同时确定枝序。枝序的确定与枝系本身发育的顺序相反, 这种方法也称为Strahler法, 也就是由外及内, 外层的第1小枝为第1级, 2个第1级相遇即为第2级, 2个第2级相遇即为第3级, 依次类推, 如有不同枝级相遇, 则汇合后取较高的枝级。计算分枝率借鉴Motomura计算式:Rb=(NN-NS)/(NT-N1), 其中, NS为最高级枝条数, NT为所有枝级中枝条的总数, N1为第1级的总枝数。逐步分枝率:Ri:i+1=Ni/Ni+1, 其中, Ni、Ni+1分别为第i和第(i+1)级的枝条总数(陈波等, 1999; 李俊清等, 2001)。
1.2.3 基因变异系数的确定2003年7月, 在A、B、C、D 4样地分别选择50株不同树龄的栓皮栎个体, 分别从每株树梢上剪下3个15 cm的嫩枝条(各个植株之间间距50 m以上), 迅速放进预先准备好的样品袋中, 然后放在冰壶中。试验时, 取每株枝条顶端上的第3个嫩叶研磨离心, 取上清液放入-20 ℃的冰箱中贮存备用。采用聚丙烯酰胺凝胶电泳法对筛选出的EST、SOD、POD、PGI、MDH、ACP、ADH和ME 8种酶系统进行分析。
2 结果与分析 2.1 栓皮栎种群萌生苗的生态学意义根据4个地区80个样方的调查资料, 对栓皮栎种群的径级分别按实生苗和萌生苗起源进行统计分析。种群径级大小按0~5 cm、5~10 cm, 每隔5 cm划分1个等级(阎桂琴等, 2001)。以径级大小为横坐标, 以个体数量为纵坐标, 做出栓皮栎种群的径级结构图, 见图 1。

|
图 1 A、B、C、D样地栓皮栎种群的径级结构 Fig. 1 Size structure of Q.variabilis population for plot A, B, C, D |
径级结构是种群的重要特征, 不同生境条件下的种群总体胸径结构特征包含在各亚种群的胸径结构信息中(刘峰等, 2000)。从实际调查的情况以及图 1中可以看出:样地A萌生苗径级在0~5、5~10、10~15 cm的个体数量占总量的14.3%, 萌生幼苗的数量占8.6%, 5~10 cm萌生的幼树占总量的2.9%, 实生幼苗个体数量占11.4%。而样地B、C和D栓皮栎萌生幼苗的数量分别占总量的4.5%、7.5%、9.3%, 实生幼苗的数量分别占总量的2.3%、5.7%、2.3%, 3个地区5~10 cm的萌生幼树分别占总数的9.1%、28.6%、8.6%。从图 1中看出, 分布中心的栓皮栎萌生幼苗幼树的数量少于实生苗的数量, 而C、D样地, 萌生苗个体数量则超过了实生幼苗的数量。由于刚萌发的实生苗的种群很不稳定, 受环境的影响很大, 从发生到死亡都有不确定性, 相比之下径级5~10 cm的实生和萌生苗处于种群的建立阶段, 对栓皮栎种群的建立、为种群幼苗后续资源的补充起了不可低估的作用。萌生的幼苗、幼树和实生的幼苗、幼树共同存在, 相互补充, 既解决了栓皮栎种群幼苗不足的矛盾, 又为种群的繁衍更新注入了新的活力。萌生作为有性生殖的一种有益的补充, 其存在大大提高了栓皮栎种群抵御外界干扰的能力。
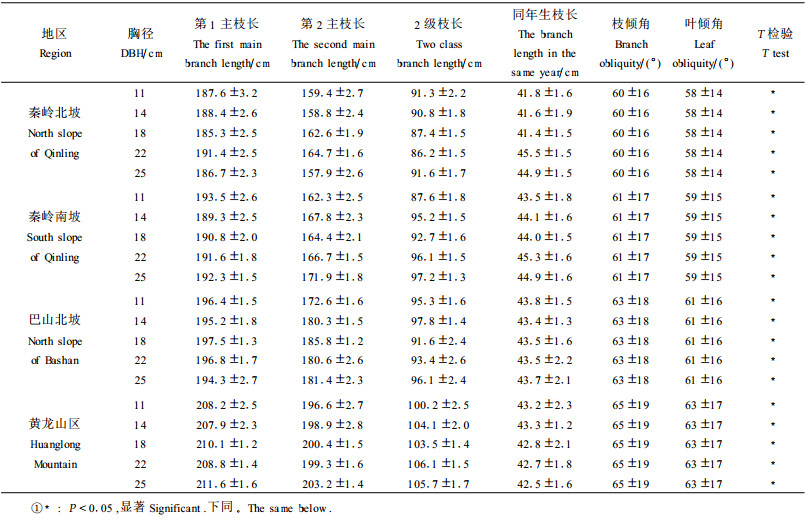
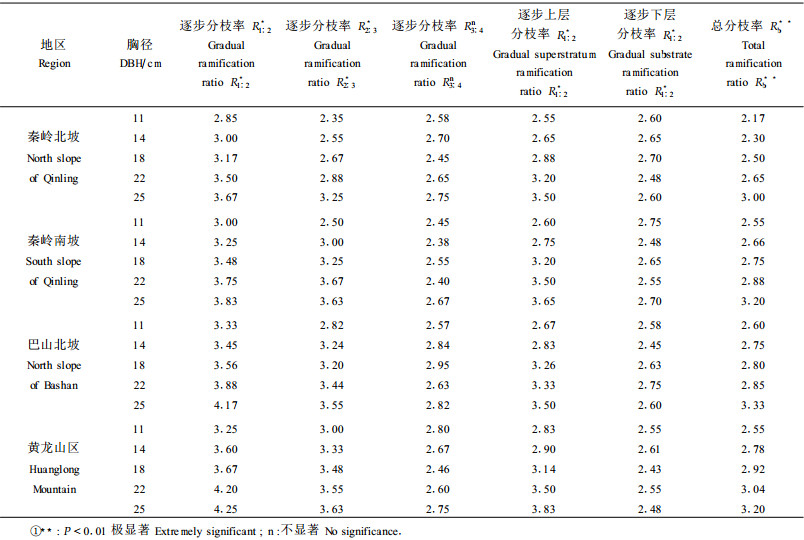
2.2 栓皮栎种群性状分化的生态学意义种群的枝长、分枝角度和分枝率等对其冠形的形成起着决定作用, 是衡量种群枝条生长、分枝能力和枝条枝级间数量配置状况的重要参数, 它们在一定程度上反映了种群的动态变化和生态特性(刘军等, 1986; Halle et al., 1978)。本文对不同研究地域栓皮栎种群的枝长、分枝角度和分枝率进行了测定, 结果见表 2和表 3。
|
|
|
|
由表 2可以看出, 每个研究地域不同胸径栓皮栎种群的1级主枝、第2主枝、3级枝条的长度基本保持不变, 而从分布中心到分布边缘当年生枝条生长速度逐渐减慢, 分枝角度呈逐渐增大趋势。在观察中我们也发现, 从分布中心到分布边缘, 栓皮栎种群的密度相应递减。在种群密度较低的地区, 枝条的顶端优势较弱, 枝条向四周辐射生长, 以充分利用生态空间, 枝条的倾角就大; 相反, 在密度大时, 为了争夺光资源以及枝条伸展的向光性, 顶端优势强, 枝条向上生长, 使倾角减小。
由表 2还可以看出, 同一栓皮栎植株的叶倾角从1级枝到3级枝逐渐增大。这种趋势使得栓皮栎种群的叶片都有一定的着生角度, 这种特有的叶片配置格局有利于叶片之间对光的折射和散射作用, 影响到冠层对光资源的利用效率。对不同地区栓皮栎的叶倾角来说, 从分布中心到分布边缘呈现出增大趋势, 说明栓皮栎种群在不同的生境中为了争取到光资源, 叶倾角的适当转向, 可以最大限度地增大叶面积来获得最大的光照资源。而枝方位角、叶方位角则是各向随机伸展的。这些性状的分化, 揭示了栓皮栎种群卓越的利用空间进行光合作用的能力, 使其能够与种内或种间的乔木种群互补利用空间和光资源, 成为群落中的优势种。
分枝率的大小能够反映栓皮栎种群的生长活力, 分枝率越大, 生长活力越强(刘峰等, 2002; Halle et al., 1978)。从表 3中可以看出, 从分布中心到分布边缘, 不同地区不同胸径的栓皮栎种群的逐步分枝率R1:2、R2:3和总体分枝率Rb表现为逐步减小的趋势。就同一地区来说, 随着径级的逐渐增大, 逐步分枝率R1: 2、R2:3呈现出增大趋势, 而R3:4呈现出无规律性, 可能是因为它反映的是栓皮栎种群过去的生长状况, 差异不明显。有些胸径小的栓皮栎种群由于分枝率小而在生态位占领方面处于劣势, 长期得不到足够的营养和光源而最终被淘汰。
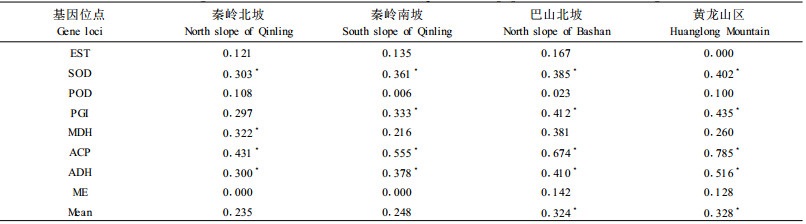
2.3 栓皮栎种群遗传变异与适应的生态学意义生境的差异反映了种群对于环境选择压力的反应, 环境选择压力的变化, 使生物适应性反应也不同(陈贤阳, 1997; Comps et al., 1991)。选择压力在地理空间上的阶梯性变化, 导致基因频率或基因分化渐变, 形成一个变异梯度(李俊清, 1995; Horn, 1991; Hamrick et al., 1992)。从表 4统计的数据看出, 酯酶同工酶(EST)、超氧化物歧化酶(SOD)的基因变异频率随生境和树龄的不同, 表现出一定的变化规律, 从分布中心到分布边缘, 这2种酶的基因变异频率呈现出逐渐减小的趋势, 而从小龄级到大龄级, 则表现出逐渐增大的趋势。过氧化物酶(POD)的基因变异频率则随着龄级的增大呈现出减小的趋势, 其他酶的基因变异频率也随着生境与龄级的不同而发生不同程度的变化。
|
|

从表 4还可以看出, 在栓皮栎分布中心的秦岭没有检测到苹果酸酶(ME)的存在, 而在过渡地带的巴山北坡和边缘的黄龙山区则检测到ME的存在; 在分布边缘的黄龙山区没有检测到酯酶同工酶(EST), 而在其他3个地区则有EST的存在。这可能是因为环境条件不同引起等位基因减少或增多, 改变了栓皮栎种群在部分分布区的遗传结构, 在一定程度上也说明了2种酶等位基因的适应意义, 揭示出植物体中的某些酶基因频率与环境变化的相关性。
另外, 根据基因的分化系数和亲近程度可以计算出不同地区栓皮栎的分化程度。从表 5可以看出, 不同地区栓皮栎的基因分化系数存在较大的差异, EST、SOD、PGI、ACP和ADH的分化系数从分布中心到分布边缘呈现出增大趋势, 即在分布中心的秦岭, 栓皮栎种群具有较高的遗传多样性, 而在分布边缘的黄龙山区, 遗传多样性降低。
|
|
综上所述, 通过等位酶分析, 可以计算不同分布区栓皮栎种群的遗传相似度和遗传距离, 进而研究栓皮栎种群的适应能力、遗传多样性和进化规律, 探索栓皮栎种群的渐变陡度与选择压力空间变化之间的关系, 从而揭示栓皮栎种群进化机制和适应对策。
3 讨论近年来, 栓皮栎受到人为破坏的现象十分严重, 所在生境的水土流失日益突出, 分布面积日益减少, 其质量和数量不断下降, 栓皮栎种群的稳定性正受到严重威胁, 研究该种群的变异和适应对策对维持种群的存在和发展具有重要的意义。从萌生苗分布情况看, 每个地区萌生苗的径级大小和数量不同, 在秦岭分布中心萌生苗的数量相对较少, 而在巴山北坡和黄龙山区, 几乎每个径级中都有一定数量的萌生苗, 尤其是低径级的数量占的比例很大。萌生幼苗在一定程度上弥补了实生苗数量的不足, 帮助栓皮栎种群度过幼苗匮乏时期, 使种群进行自我更新。因此, 萌生苗作为有性生殖的一种有益补充, 是栓皮栎种群应对外界环境变化的一种适应性对策, 这种适应是进化性的, 它的存在大大提高了栓皮栎种群对环境变化的反应速度和适应能力, 对于维持栓皮栎种群的整体活力、遗传多样性、种群的稳定性都具有十分重要的生态学意义。
为适应不同的生境条件, 栓皮栎种群的枝长、分枝角度和分枝率等性状参数也发生了变异。从分布中心的秦岭到分布边缘的黄龙山区, 当年生枝条的长度和逐步分枝率R1:2 、R2:3以及总体分枝率Rb均呈逐渐减小的趋势, 而分枝角度和叶倾角则逐渐增大。栓皮栎通过形态上的可塑性来完成对生境异质性的反应, 形成与环境相适应的有利特征, 使不同生境中栓皮栎的枝长、枝倾角和分枝率处于最佳组合, 以保证最高的光合效率, 这是栓皮栎种群的一种适应性进化对策。
生境的差异使栓皮栎外观形态表现不同的同时, 在微观上也表现出基因频率或基因分化的渐变, 形成一个变异梯度。在栓皮栎生长期内, 随着生境条件的不同, 栓皮栎中各种酶的基因变化频率也在升高或下降。EST、SOD的基因变异频率随生境和树龄的不同, 表现出一定的变化规律, 即从分布中心到分布边缘, 这2种酶的基因变异频率呈现出逐渐减小的趋势; 从小龄级到大龄级, 则表现出逐渐增大的趋势, 而过氧化物酶(POD)则随着龄级的增大呈现出减小的趋势。同时, 不同生境条件下栓皮栎的基因分化系数存在较大的差异, EST、SOD、PGI、ACP和ADH的分化系数从分布中心到分布边缘呈现出增大趋势, 揭示出栓皮栎中的某些酶的基因频率和遗传分化与环境变化相关。尽管本研究中一些等位基因频率的变化与生态条件未表现出相关性, 但从已有的文献来看, 种群的生态学和生活史特征与某些等位酶的变异性水平和分布之间确实存在着相关性(李丹等, 2001; Hamrich et al., 1992)。
从以上的分析可以看出, 陕西地区的栓皮栎种群由于处在不同的生境条件下, 种群之间存在着一定的生境梯度, 受到气候、土壤、地形、生物和人为等不同因素影响, 栓皮栎种群在数量、外部形态多样性、适应性性状分化和遗传结构等方面都发生了很大的变化。包括径级结构、树冠构件型的变化以及基因频率随环境梯度的变化, 都是栓皮栎种群动态的表现形式, 是栓皮栎种群在不同生境条件下与环境长期适应和进化的结果。自然选择的压力就是种群进化的动力, 这种动力的外观表现就是栓皮栎种群的动态的变化, 种群的动态实际上就是一种适应。因此, 在种群水平上, 从宏观和微观的角度, 运用定性和定量、室内和室外相结合的方法, 将生境因子、表型变异的数量和分子动态的基因频率有机地结合起来, 揭示了栓皮栎种群变异和适应策略, 为种群动态与分子动态机制的研究奠定了基础。
陈贤阳. 1997. 西南桦种群的基因多样性和分化. 植物学报, 39: 149-155. |
陈波, 宋永昌. 1999. 木本植物的构型及其在植物生态中的作用. 生态学报, 19(3): 359-364. DOI:10.3321/j.issn:1000-0933.1999.03.012 |
李俊清, 臧润国. 2001. 欧洲水青冈(Fagus sylvatical L.)构筑型与形态多样性研究. 生态学报, 21(1): 152-155. |
李俊清. 1995. 中国亚热带地区海滩森林稀有和濒危物种的基因多样性保护和开发. 北京林业大学学报, 4: 1-16. |
刘峰, 陈伟烈. 2002. 神农架地区辽东栎种群的结构和更新. 生态学报, 24(4): 396-401. DOI:10.3321/j.issn:1005-264X.2002.04.003 |
刘军, 齐国良. 1986. 天然红松种群的波动特征. 生态学杂志, 5(5): 1-5. |
李丹, 彭少麟. 2001. 三个不同海拔梯度马尾松种群的遗传多样性及其与生态因子的相关性. 生态学报, 21(3): 415-421. DOI:10.3321/j.issn:1000-0933.2001.03.011 |
吴明作. 1998. 栓皮栎研究进展. 陕西林业科技, 4: 65-69. |
闫桂琴, 赵桂仿. 2001. 秦岭红杉种结构和动态研究. 应用生态学报, 12(6): 824-828. DOI:10.3321/j.issn:1001-9332.2001.06.006 |
袁嘉祖, 张汉雄. 1991. 黄土高原地区森林植被建设的优化模型. 北京: 科学出版社.
|
中国树木志编委会. 1976.中国树木志.北京: 中国林业出版社, 12: 507-515
|
Halle F, Oldeman R A, Tomlinson P B. 1978. Tropical trees and forests—an architectural analysis. Berlin:Heidelberg: Springer-Verlag.
|
Comps B, Petit R J. 1991. Genetic variance of the Croatian beech tands. Ann Sci For, 48: 15-28. DOI:10.1051/forest:19910102 |
Hamrick, Godt J L. 1992. Factors influencing levels of genetic diversity in woody plant species. New Forest, 6: 95-124. DOI:10.1007/BF00120641 |
Ho rn. 1991. Canopy disturbance patterns and regeneration of Quercus species in two Ohio old growth forests. Vegetatio, 93: 9-18. |