 2005, Vol. 41
2005, Vol. 41文章信息
- 王喜明, 于建芳, 苏金梅, 李瑞英.
- Wang Ximing, Yu Jianfang, Su Jinmei, Li Ruiying.
- 木材干燥应力应变超微观模型的构筑
- The Development of Ultrastructural Model on Wood Drying Stress-Strain
- 林业科学, 2005, 41(4): 204-209.
- Scientia Silvae Sinicae, 2005, 41(4): 204-209.
-
文章历史
- 收稿日期:2003-06-10
-
作者相关文章


木材干燥应力应变研究不论在理论上探索还是用于指导生产实践,都具有重大的意义。然而木材干燥应力与应变的关系是非常复杂的, 因为木材属于既有弹性又有塑性的材料——弹塑性材料。应变的大小受很多因素的影响,包括材料自身的因素,例如密度、管胞或纤维胞壁的微纤丝的角度以及恒定或变化的环境因素(如大气的温度、相对湿度)等。还有依从于时间的因素(申宗圻, 1990)。正是由于木材本身的特殊结构及其在干燥过程中各参数变化的复杂性,建立一个精确性较高和适用性较广的干燥应力应变数学模型很困难。若不靠数学物理方法,单纯的试验很难重现实际的干燥过程。理论模型体现了干燥过程中各参数间的定量关系和规律,用试验数据验证理论模型的正确性及适用性很重要,然而,即使模型得不到广泛的试验证实,也可借助计算机模拟,不仅可以定性地更好地理解干燥过程,而且可以定性地考察各参数之间的依存关系和变化趋势。所以,建立理论模型对干燥工艺进行定性地分析研究,是很有意义的。
在探讨木材干燥中应力应变的发展变化时,有必要从细胞壁的微观构造出发建立细胞结构模型, 弄清细胞壁层每一成分的性能。在木材细胞壁模型化中,最重要的结构参数是纤维素、半纤维素和木素在不同壁层中的数量与分布,在不同壁层上微纤丝的排列角度以及壁层厚度;最重要的物性参数是纤维素、半纤维素和木素的弹性常数和应变特性。而且建模时所用的参数值一般是建立在文献基础上,一些可由试验确定,一些可由理论估计,还有一些是基于其他材料所作的假设。
1 建模基础一般地,木材细胞壁可看成是由不同层组成的层合结构,一些研究者利用经典的层合板理论,把木材细胞壁看成是复合层来模拟木材的收缩和各向异性。一般运用层合理论对细胞壁模型进行计算时的基本步骤是:首先分析组成模型各层的微观力学性质;其次运用层合理论把这些性质结合起来,估计整个细胞壁结构的性质(Lennart et al., 1985)。用这种方法也可描述由于含水率变化引起的尺寸改变。正常木材细胞壁由胞间层、初生壁和次生壁组成,每一层都由于其化学组成、厚度及纤丝角不同而在木材细胞壁收缩中扮演着重要角色,在理论上已经提出几个模型来描述细胞壁的收缩特性。例如,Innes(1995)建立了木纤维细胞应力模型, 预测了应力和应变在细胞壁上的分布。Barber等(1964)依据木材细胞的微观构造, 认为管胞是由S2层组成的矩形壳,并运用层合理论分析了细胞壁的收缩,认为木材总的收缩应变是纤丝角的函数。Barber(1968)假设木材细胞壁是厚壁圆柱状, 推导出应变比和纤丝角的关系,此模型和木纤维的形状相似, 不仅能预测木纤维长度的改变, 而且可以预测木纤维直径的改变。各国学者在Barber模型的基础上,对其假设作不同程度的修改,分别发展成各自的细胞壁模型,从不同角度分析研究细胞壁的收缩特性及影响其各向异性收缩的因素。Yamamoto(2002)发展了多层木纤维模型,从理论上讨论了应力应变关系,预测了木纤维收缩膨胀的各向异性,其中把含水率的变化作为显式参数;但他假设微纤丝在横向所有的剪切模量、泊松比和弹性模量均为零。由于在干燥过程中,沿纤丝方向发生的应变比沿横向的要小,而且S2层占整个细胞壁很大比例,所以本文着重探讨细胞壁S2层在失水时的应力应变发展规律,分析含水率和微纤丝角的影响。
2 模型的构筑细胞壁的主要成分是纤维素、半纤维素和木素,这些成分的物理性质在很大程度上依赖于含水率,含水率很高时胞壁变软,含水率较低时变硬,而且不同的试样失水不同,导致应变也不同。此外,微纤丝在不同部位的排列方向不同,在干燥中对不同方向所产生的应变也有较大的影响,所以,本文建立超微观模型时考虑含水率与微纤丝角这2个显式参数。
假设S2层由2部分组成,即骨架和基体。骨架是纤维素分子聚集而成的微纤丝,基体是半纤维素和木素的聚合物。S2层模型图如图 1(Barber, 1968)。

|
图 1 S2层模型图 Fig. 1 A model of S2 layer |
建模时所作的假设(Yamamoto, 2002):1)基体的应变是各向同性的,且只发生正应变,骨架的应变是各向异性的;2)木纤维模型收缩引起细胞壁每一点的位移是连续的;3)纵向位移不依赖于半径r;4)剪应变γLT=0;5)S2层体积不可压缩。
2.1 基体的应力应变关系应用弹性理论并结合假设1)可推出基体的应力应变关系式(Barber et al., 1964)为:

|
(1) |
式中:L、T、R表示细胞纵向、弦向和径向;m代表基体;ε2表示S2层基体的收缩应变;K表示体积弹性模量。S表示剪切模量。计算公式如下:
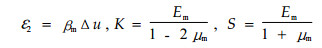
|
(2) |
式中:βm是基体的线干缩系数;Δu是由一种状态到另一种时含水率的变化量;Em和μm分别是S2层基体的杨氏模量和泊松比。
2.2 骨架应力应变关系图 2(Yamamoto, 2002)表示纤维素骨架的一个平面小微元体。假设L、T、R这3个方向正交;x表示与纤维素分子链平行的方向,如图建立局部坐标系,z轴方向和细胞径向R方向一致,θ表示微纤丝角。

|
图 2 骨架微元体 Fig. 2 An element of CMF |
由于微纤丝包括非结晶区,所以失水在一定程度上会引起微纤丝产生应变,而且细胞失水时,由于受到纤维素结晶区的抑制,沿微纤丝长度方向的应变相比沿横向的应变要小得多,因此本文只考虑沿微纤丝横向的收缩应变,记为εs,认为木材含水率从纤维饱和点到全干时,εs随含水率线性变化(李维桔, 1983),且细胞壁层的含水率是相同的,所以有:
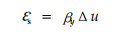
|
(3) |
其中:βy是微纤丝在横向的线干缩系数。
在局部坐标系xyz中,由广义虎克定律可写出S2层微纤丝的应力应变关系式:

|
(4) |
只考虑x和y方向上的线弹性应变,则有:Q11=e1, Q22=E2其余Qij=0, E1和E2分别是x和y方向上的弹性模量,所以上式简化为:

|
(5) |
转换坐标系可推出在LTR坐标系中骨架的应力应变关系式:
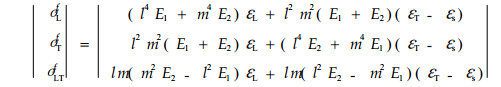
|
(6) |
其中:l=cosθ, m=sinθ, θ是微纤丝角。
2.3 S2层应力应变关系由叠加原理知,作用在细胞壁任何单元上的应力可以认为是基体应力与骨架应力之和,所以有:
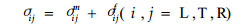
|
(7) |
这样,可推得S2层总应力应变关系式为:
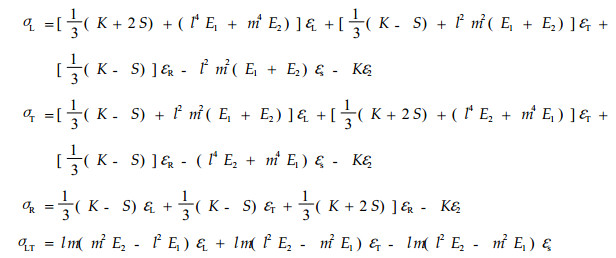
|
(8) |
考虑形变协调关系:
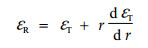
|
(9) |
则(8)式转化为:
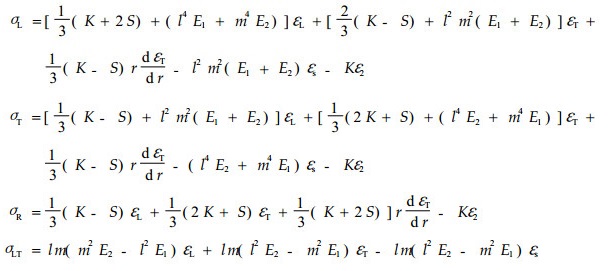
|
(10) |
这就是细胞壁S2层的应力应变关系式。由假设5)可认为μm =0.5(徐秉业等, 1986),这样K就远远大于E1、E2、S,即可认为E1/K=E2/K=S/K =0,并把上式看成是εL、εT和rdεT/dr的线性非奇次方程,则可导出εL的表达式:
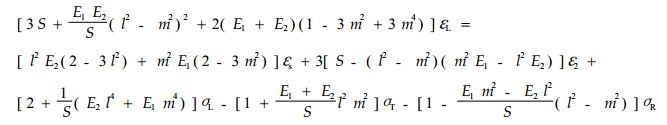
|
(11) |
结合S2层的边界条件,纵向应力::

|
(12) |
径向应力
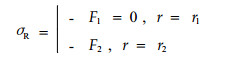
|
(13) |
可推得εL的表达式:

|
(14) |
同理可求出εT的表达式:

|
(15) |
(14) 和(15)式中的系数如下:
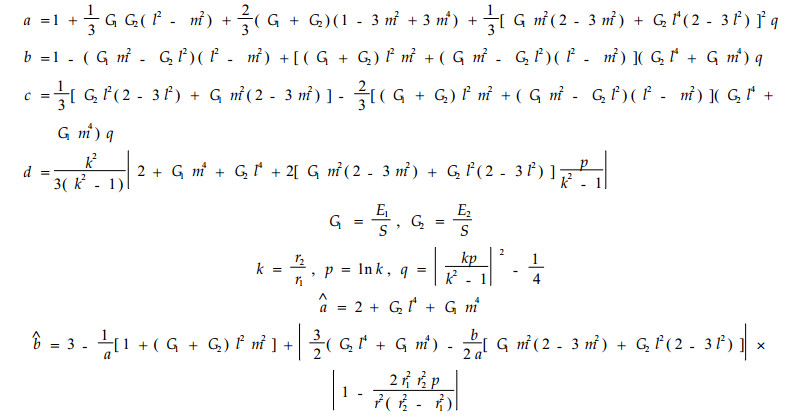
|
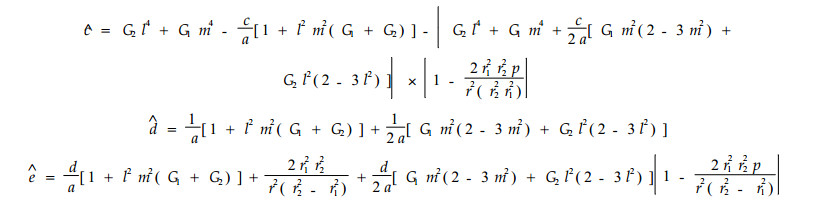
|
一般地,理论数学模型都是建立在一定的假设基础上,模型中所含的物性参数和几何参数带有普遍意义。对木材来说,它是一种各向异性的弹塑性材料,不同的树种,甚至同一树种的不同部位、不同方向,在干燥过程中产生的应力与应变都有很大的不同。由于试验条件的限制,本文只选取一些参数值对理论模型作预测。本文对弦纵向应变模型预测所用的参数(Innes, 1995;Barber et al., 1964)如下:

|
F0,F2,r1,r2的值参考Inners(1995),其余参数的值是参考Barber等(1964)。

|
图 3 应变随微纤丝角变化曲线 Fig. 3 Strain in longitudinal and tangential directions vs MFA |

|
图 4 应变随含水率变化曲线 Fig. 4 Strain in longitudinal and tangential directions vs moisture content |
微纤丝角等于零时,纵向应变远小于弦向应变,随微纤丝角的增加纵向应变增加而弦向应变减小,这说明微纤丝角对木材应变有一定的影响。
在微纤丝角小于0.4弧度的区域,纵向应变较小,弦向应变较大,但变化都较慢,纵向应变增加约0.02个百分点,弦向应变减小约0.2个百分点。
当微纤丝角大于0.4弧度时,弦纵向应变梯度都在增加,微纤丝角增加0.28弧度,纵向应变就增加约2个百分点,弦向应变减少约3个百分点, 这主要是微纤丝对横纵向应变的抑制作用。
随微纤丝角的增加,弦向应变减小的幅度比纵向应变增加的幅度要大,在约0.68弧度时弦纵向应变相等,约达到2.6%,在大于0.68弧度的区域,纵向应变反而大于弦向应变,在0.7弧度时纵向应变高达3.4%,而弦向应变降到1.6%。这充分说明微纤丝角对木材弦纵向应变的影响很大。
含水率达30%时,纵向应变大于零,弦向应变等于零,且随含水率的降低弦纵向应变逐渐增大,直至含水率降到零应变达到最大为止,二者成线性变化关系。
含水率从30%降到0,弦向应变增加8%~9%,纵向应变增加却不到1%,很明显弦向应变大于纵向应变。此外,在含水率变化过程中,应变差异也在增加。
4 小结由于从细胞壁的微观构造出发建立细胞结构模型,更有助于理解干燥的物理过程,从微观的角度解释干燥应力应变的发展规律,所以很多学者在这方面做了研究,他们从不同的角度作相应的假设,分别给出了不同的细胞壁结构模型,有方形的,圆形的,两层的,三层的,甚至七层的。尽管所建的模型不同,但是对指导实际干燥工艺的意义是相似的,而且有很多模型预测结果是相似的,这正体现了建立理论模型的重要意义。本文建立的干燥应力应变模型,适用于从微观角度解释木材的干燥特性。模型预测结果与木材干燥理论基本一致,与相关文献的结果也相似,为干燥工艺提供了一定的理论参考。根据木材超微观结构特点,利用数学和力学原理,采用数学建模与计算机辅助相结合的研究方法,建立的关于细胞壁S2层的应力应变模型为:

|
推导出应变的变化规律为:
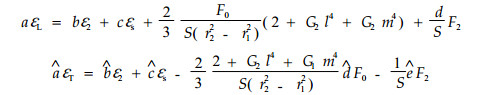
|
建立模型时假设S2层是由骨架和基体组成,骨架是纤维素分子聚集而成的微纤丝,基体是半纤维素和木素的聚合物,基体变形是各向同性的,骨架的变形是各向异性的,而且纵向应变不依赖于半径r,S2层体积是不可压缩的。所建模型的特点是把含水率和纤丝角作为显式参数,在基本参数取定的前提下,由数学软件绘图可以明显地看出改变含水率和纤丝角对弦纵向应变的影响,且含水率和弦纵向应变是线性关系。在试样纵向相比弦径向较长时,所建模型适用于从微观角度分析整个干燥过程中应力应变的发展规律。
李维桔. 1983. 木材弹性及木材干燥应力Ⅱ:木材干燥应力. 南京林产工业学院学报, 2: 107-122. |
徐秉业, 陈森灿. 1986. 塑性力学简明教程. 北京: 清华大学出版社.
|
申宗圻. 1990. 木材学. 北京: 中国林业出版社.
|
Barber N F. 1968. A theoretical model of shrinkage wood. Holzforschung, 3: 97-103. |
Barber N F, Meylan B A. 1964. The anisotropic shrinkage of wood. Holzforschung, 18: 146-156. DOI:10.1515/hfsg.1964.18.5.146 |
Innes T C. 1995. Stress model of a wood fiber in relation to collapse. Wood Science and Technology, 29(5): 363-376. |
Lennart S, Alf de R. 1985. A model for the prediction of fiber elasticity. Wood and Fiber Science, 17(3): 336-350. |
Yamamoto H. 2002. Predicting anisotropic shrinkage of softwood. part Ⅰ:theories. Wood Science and Technology, 36: 75-91. DOI:10.1007/s00226-001-0122-4 |