 2005, Vol. 41
2005, Vol. 41文章信息
- 张冬梅, 鲍甫成, 张志毅, 黄荣凤.
- Zhang Dongmei, Bao Fucheng, Zhang Zhiyi, Huang Rongfeng.
- 毛白杨无性系湿心材比例的遗传分析
- Genetic Analysis of Wetwood Proportion on Clone Test Stand of Populus tomentosa
- 林业科学, 2005, 41(4): 140-144.
- Scientia Silvae Sinicae, 2005, 41(4): 140-144.
-
文章历史
- 收稿日期:2003-03-24
-
作者相关文章


2. 中国林业科学研究院木材工业研究所 北京 100091;
3. 北京林业大学 北京 100083
2. Research Institute of Wood Industry, CAF Beijing 100091;
3. Beijing Forestry University Beijing 100083
湿心材是树木生长中的一种反常现象,是树木加工利用中的一大难题(祖勃荪,2000)。Ward(1986)总结称,21种针叶材中13种有湿心材,50种阔叶材中38种有湿心材。国外对杨树湿心材的系统研究始于20世纪50年代,包括杨树湿心材的形成,湿心材形成与细菌、气体和水分的关系,湿心材的酸度,材色,湿心材的力学强度及渗透性等。结果表明:与正常材相比,由于湿心材含水率高、颜色深、抽提物多以及pH值偏酸或偏碱性,使木材物理、化学性质发生变化,导致干燥时易发生皱缩及沿年轮开裂、刨削难,呈碱性的湿心材很难用脲醛树脂胶合以及因变色导致的商品价值大幅下降等加工问题,直接影响到杨木木材的干燥、制材和胶合板的质量(Clausen et al., 1952;Kemp,1959;Haygreen et al., 1966)。
我国湿心材的研究较晚。杨树是我国制浆造纸业的主要树种,由于湿心材现象的普遍发生,致使湿心材的综纤维素含量和α-纤维素含量比正常材低,直接影响到出浆率以及印刷光泽度,这对制浆造纸业极为不利(安培均,1992;姜笑梅等,1993;诸葛强等,1997;晁龙军等,1996;1997;1998)。本文研究毛白杨(Populus tomentosa)湿心材所占比例,并对其进行遗传分析,为选育出无湿心材或湿心材比例低和木质素含量低的优良纸浆材毛白杨无性系提供科学依据。
1 材料与方法 1.1 材料试材取自于1987年定植于山东冠县苗圃的纸浆材毛白杨无性系测定林。测定林中的无性系是北京林业大学毛白杨协作组从毛白杨分布区范围内选优得到的(朱之悌,1992)。株行距4 m×5 m,8次重复。
1.2 方法选取测定林中的25个无性系各6株,用9 mm生长锥在树木胸径处分南北向钻取木芯(弃去有疤木、应拉木及其他木材缺陷的木芯)。量取湿心材长度占整个木芯的比例,并测量取样单株的树高、胸径和材积。
截取不同无性系单株木芯上第9年轮的木材试样,采用最大含水量方法对木材试样的基本密度进行测定。将每个无性系6个单株的的木芯混合(包括湿心材和非湿心材部分)、干燥、粉碎、制成木粉,测定不同无性系间木质素含量。木质素含量测定方法依照GB 2677.8-81进行。
木材白度利用MSC-P多光源分光测色仪测定, 属于甘茨白度值。每个无性系6个单株的试样各测6个点,然后取平均值。计算公式采用我国用于评定纺织品白度方法(GB 8425-87)中规定用的D65光源、1964 10°标准观察者数据,公式为:W10 =Y10+ 800(0.313 8-x10)+1 700 (0.3310-y10)。
2 结果、分析与讨论 2.1 毛白杨无性系湿心材的遗传变异改良木材的一条有效途径就是培育出具有所希望木材性质的树木品系。大多数木材的性质具有中等至较高的遗传性,因此通过遗传手段可以使其向我们需要的方向变化,木材的湿心材也不例外。
杨树湿心材的遗传变异较为复杂,从150个木芯试样的湿心材形态看,无性系间以及无性系内单株间的湿心材比例表现不一。从表 1可清楚地看出,毛白杨无性系间湿心材百分比存在着明显的差异。湿心材比例较高的无性系有40#、1210#、1808#、11#、3332#和1 005#无性系,它们的湿心材比例分别达32.9%、27.3%、26.8%、26.1%、25.6%和24.1%;而有的较低,如:41#、27#、3-2-1#、60#和5101#无性系的湿心材比例分别为5.5%、7. 5%、8.2%、8.8%和9.8%。无性系内单株间湿心材比例的变异幅度和变异系数也表现出很大的差异,如34#、3-1-27#、5101#、60#、3-23-32#、27#无性系内单株间变异幅度较大,变异系数分别为122.68%、119.69%、100.64%、110.39%、90.33%、和89.40%;而1808#、11#、1232#和66#无性系的湿心材比例在单株间的变异较小,变异系数分别为12.52%、18.01%、20.9 5%和29.55%。
|
|
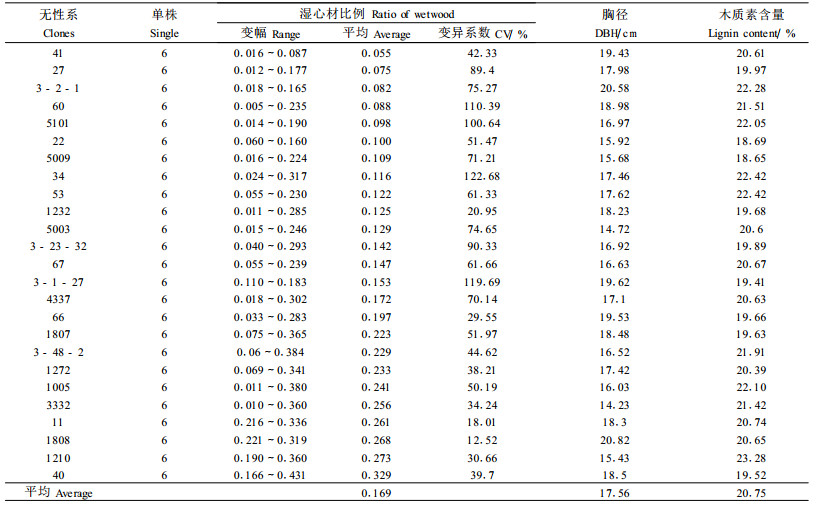
对湿心材比例进行方差分析见表 2。无性系南北木芯方向间的湿心材比例差异不显著,无性系间湿心材比例差异达到5%显著水平,无性系内湿心材比例在5%水平上差异不显著。变异是选择的基础,毛白杨无性系间湿心材比例差异的显著性为选择性育种奠定了基础。对毛白杨无性系木材湿心材比例的遗传参数估计结果(表 2)表明,无性系间湿心材比例的无性系重复力高达0.749,受中度遗传控制;无性系内的湿心材比例个体重复力为0.323,受弱度遗传控制。从而可推测不同杨树无性系的湿心材产生主要是受遗传控制的,有望通过无性系选择获得湿心材比例低的无性系。美洲黑杨(Populus deltoides)新无性系的湿心材遗传力高达0.689 1,表明美洲黑杨湿心材性状变异也主要由无性系和基因型控制的(诸葛强等,1997)。
|
|
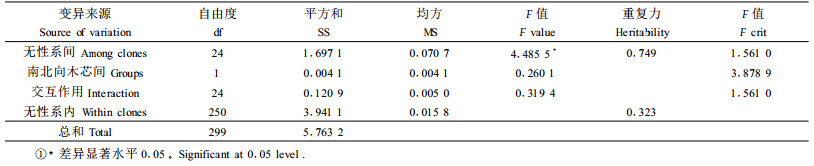
有研究报道湿心材的形成分2种:原发性湿心材和继发性湿心材。原发性湿心材是指细菌在幼苗繁殖时已经潜伏于苗木中,随着树木的生长,细菌开始活动,逐渐形成湿心材。继发性湿心材则是由于苗木起初没有细菌,在生长过程中因受到生物或非生物的伤害,细菌由伤口进入树木形成层等诱发湿心材发生(祖勃荪,2000)。从本研究结果看,毛白杨木材无性系间的湿心材含量受中度的遗传控制,说明该性状与杨树本身固有的生理因素及结构有关,受树木生长后期的环境因子的影响相对较小。推测毛白杨木材的湿心材形成属于原发性湿心材,可能是由寄生菌造成的病原后果,原因也是毛白杨本身的生理特征及结构适合于细菌潜伏于繁殖材料中,随着树木的生长,细菌开始活动,逐渐形成湿心材。对于毛白杨木材的湿心材的形成还需对病菌进行分离、纯化,导入健康植株等进一步的研究。
2.2 毛白杨无性系湿心材比例与木材白度的关系木材白度即木材本身的光散射和反射性能,又称木材色度。木材本身的颜色是影响漂白后纸浆白度的重要因子。研究发现,纸浆白度与所用木材白度有良好的相关性,而且纸浆可漂性与木材初始白度亦有一定关系(房桂干等,1995)。姚光裕(1997)认为高白度杨木化学浆是生产高级纸张的一种重要配比浆原料。
通过对毛白杨无性系木芯试样颜色特征(人为分为白和偏红两类)的观察和白度的测定发现,25个无性系中9个无性系的木芯试样呈现较白的外部特征(表 3),其中8个无性系的白度测定值大于平均数,即3-48-2#、3-2-1#、4337#、41#、22#、5101#、66#和1232 #无性系。而这些无性系中湿心材比例低于平均水平的有41#、3-2-1#、5101#、22#和1232#无性系。显然,在湿心材比例和木材白度这2个性状上可选择这些无性系。然而,选择时应结合每个无性系湿心材比例的变异系数(表 1)综合分析,在这几个无性系中1232#的单株变异系数较小为20.95%,说明该无性系在湿心材比例这个性状上受外界因素的影响较小,容易遗传控制,显然,1232#是材质较白湿心材比例较低且能稳定遗传的无性系。41#无性系的变异系数为42.33%,也可以考虑。尽管3-2-1#、22#和5101#无性系的湿心材比例较低,但这些无性系单株的湿心材比例变异较大,受遗传外的因素影响大些,显然不是最佳选择。
|
|
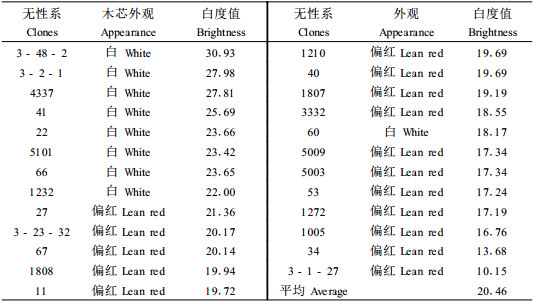
根据文献记载,湿心材的形成与细菌有关,由于细菌的活动使湿心材的构造、化学、物理力学性质发生变化(Schink et al., 1981; Rink et al., 1987;1989;Yanig et al., 1989; Wilkins et al., 1990)。也有报导认为湿心材的形成和树木细胞产生的多种酶如多酚氧化酶、过氧化物酶、纤维素酶、淀粉酶、苯丙氨酸酶等生理活动相联系(Loewas, 1976; Yanchuk et al., 1986)。胡景江等(1999)研究发现,木质素是植物体内的一种重要的物理抗菌物质,它能与羟脯氨酸糖蛋白(HRGP)一起作为结构屏障物,起强固细胞壁的作用,认为HRGP和木质素与杨树对溃疡病的抗性有关。本文将25个无性系的每个无性系的6个单株湿心材比例的平均数和相对应的6个单株的无性系木质素含量平均数进行了相关分析,结果表明,二者相关系数为0.199 1(F0.05= 0.381),未达到相关显著水平。显然,对湿心材的比例和木质素含量这2个性状可以独立进行选择。在木质素含量性状上,低于25个无性系总平均水平(20.75)的有16个无性系(表 1),可供选择。结合木芯试样白度测定值(表 3),16个无性系中4337#、41#、22#、5101#和1232#无性系的白度测定值高于平均数,因此,欲选择木质素含量低、色较白的树种时可选择这些无性系。
本次试验的试材是木芯,而木质素分析需要的木粉量较大,所以木质素含量只有利用每个无性系6个单株的木芯混合物测定,从而造成湿心材比例和木质素含量的相关分析是无性系的单株平均数间的结果。毛白杨树木个体内,正常材木质素含量和湿心材木质素含量的变异情况以及湿心材比例与湿心材处木质素含量的关系,还需进一步探讨。
2.4 毛白杨无性系湿心材比例与密度及木材生长性状关系为了进一步揭示毛白杨无性系湿心材比例与木材密度及生长性状的关系,将毛白杨无性系测定林中的25个无性系的150单株的基本密度(ρ)、胸径(D)、树高(H)和材积(V)指标,与无性系湿心材比例之间做简单相关分析,并作散点图(图 1 A~D)。图中横坐标为各无性系的湿心材比例(W),纵坐标分别为无性系的基本密度、胸径、树高及材积。毛白杨无性系湿心材比例分别与木材的基本密度、胸径、树高及材积呈负相关关系,表型相关系数分别为-0.019 9、-0.374 3**(F0.01= 0.208)、-0.155 8和-0.120 2,其中湿心材比例与胸径在1%的水平上呈现显著的负表型相关性。

|
图 1 毛白杨无性系湿心材比例与树高、胸径、材积及基本密度的散点图 Fig. 1 Scattered diagrams of relationship between wetwood proportion(W) and growth traits(H, D, V) and basic density(ρ) A基本密度Basic density, B胸径DBH, C树高Height, D材积Volume. —纵坐标性状的数据平均值The average of property on abscissa; |横坐标性状的数据平均值The average of property on ordinate. |
生物性状的变异来源于2个方面,一是遗传因素决定的,称遗传变异;二是受外界条件影响而产生的,称环境变异。遗传因素和环境因素的联合作用决定了生物性状的表型变异。对林木育种来说,性状间的遗传相关对遗传改良意义更大。通过对毛白杨无性系湿心材比例和木材的密度、胸径、树高及材积的遗传相关分析,遗传相关系数分别为:0.000 0、-0.425 8** (F0.01= 0.208)、-0.123 4和-0.160 2*(F0.05= 0.159)。结果表明湿心材比例分别与木材基本密度以及树高之间的遗传相关性都很微弱,说明湿心材比例和木材基本密度、树高分别受相互独立的遗传基因控制,它们之间可以独立选择;湿心材比例与材积、与木材胸径之间分别在5%和1%的水平上呈显著的负遗传相关。推测毛白杨无性系湿心材的形成有可能与生长过程中胸径、材积的增加受到连锁基因的控制作用。
有研究认为有湿心材的树可以继续活几十年,对木材强度也不造成严重损害(祖勃荪,2000)。本研究结果显示湿心材比例与木材胸径以及湿心材比例与材积的遗传负相关达到显著水平,所以在湿心材比例低于平均水平且胸径较大的41#、27#、3-2-1#、60#、53#、1232#无性系中,可选择湿心材比例变异系数较小的41#和1232#无性系。
3 结论毛白杨无性系间湿心材比例差异达到5%显著水平,无性系重复力高达0.749,湿心材的产生主要是受遗传控制,无性系间选择有效,有望获得无湿心材或湿心材比例较低的优良毛白杨无性系。
毛白杨湿心材比例与木质素含量及与木材基本密度之间的相关系数分别为0.199 1和-0.019 9,相关不显著,这3个性状可以独立进行选择;湿心材比例与胸径在1 %水平上达到极显著负表型相关和极显著负遗传相关,可联合选择。
在所研究的25个毛白杨无性系中,可选择1232#无性系,它的湿心材比例较低、木质素含量较低、材色较白,且胸径较大。
安培均. 1992. 毛白杨红心病对材性和利用的影响. 陕西林业科技, (1~2): 46-53. |
晁龙军, 朴春根, 曾大鹏. 1996. 杨树湿心材发生及危害的调查研究. 林业科学研究, 9(Mem): 121-124. |
晁龙军, 龚益鸿, 郭先梅, 等. 1997. 湖北省杨树湿心材发生及危害的调查研究. 湖北林业科技: 13-15. |
晁龙军, 曾大鹏, 孙福在, 等. 1998. 引起杨树湿心材的一种病原真菌——Fusarium proliferatum(Matsushima)Nirenberg. 林业科学, 34(5): 69-73. DOI:10.3321/j.issn:1001-7488.1998.05.009 |
房桂干, |
胡景江, 朱玮, 文建雷. 1999. 杨树细胞壁HRGP和木质素的诱导积累与其对溃疡病抗性的关系. 植物病理学报, 29(2): 151-156. DOI:10.3321/j.issn:0412-0914.1999.02.011 |
姜笑梅, 张立非, 徐邦兴, 等. 1993. I-69杨湿心材发生、分布及材性的研究. 林业科学研究, 6(5): 480-485. |
姚光裕. 1997. 杨木纸浆技术的新进展. 世界林业研究, 2: 44-49. |
朱之悌. 1992. 全国毛白杨优树资源收集、保存和利用地研究. 北京林业大学学报, 增(3): 1-25. |
诸葛强, 黄敏仁, 潘惠新, 等. 1997. 杨树湿心材的化学特性及形成机理研究. 林业科学, 33(3): 259-266. |
祖勃荪. 2000. 国外对杨树湿心材的研究. 林业科学, 36(5): 85-91. DOI:10.3321/j.issn:1001-7488.2000.05.015 |
Clausen V H, Kauferr F H. 1952. Occurrence and probable cause of heartwood degradation in commercial species of Populus. F P J, 2(4): 62-67. |
Haygreen J G, Wang S S. 1966. Some mechanical properties of aspen wetwood. F P J, 16(9): 118-119. |
Kemp A K. 1959. Factors associated with the development of collapse in aspen during kiln drying. F P J, 9(3): 124-130. |
Loewas F A. 1976. The structure, biosynthesis and degradation of wood. Rescent Advances in Phytochemistry, 11: 247-290. |
Rink G, Phelps J E. 1989. Variation in heartwood and sapwood properties among 10-year-old black walnut trees. Wood and Fiber Science, 21(2): 177-182. |
Rink G. 1987. Heartwood color and quanlity variation in a young black walnut progeny test. Wood and Fiber Science, 19(1): 93-100. |
Schink B, Ward J C, Zeikus J G. 1981. Microbiology of wetwood:importance of pectin degradation and clostridium species in living tress. Applied and Environment Microbiology, 42(3): 526-532. |
Ward J C. 1986. The effect of wetwood on lumber drying times and rates:an exploratory evaluation with longitudinal gas permeability. Wood and Fiber Science, 18(2): 288-307. |
Wilkins A P, Stamp C M. 1990. Relationship between wood color silvicultural treatment and rate of growth in Eucalyptus grandis Hill (Maiden). Wood Sci Technol, 24: 287-304. |
Yanchuk A D, Micko M M. 1986. Genetic variation of extractives in the wood trembling aspen. Wood Sci Technol, 22: 67-71. |
Yanig K C, Hazernberg G. 1989. Relationship between tree age and sapwood/heart wood width in Populus tremuloidis Michro. Wood and Fiber Science, 21(2): 177-182. |