 2005, Vol. 41
2005, Vol. 41文章信息
- 刘震, 毕会涛, 蒋建平, 王联营, 乔保水.
- Liu Zhen, Bi Huitao, Jiang Jianping, Wang Lianying, Qiao Baoshui.
- 泡桐侧芽萌发成枝接干规律
- The Laws of Branching and Trunk Extension Sprouted from Lateral Buds in Paulownia trees
- 林业科学, 2005, 41(4): 42-47.
- Scientia Silvae Sinicae, 2005, 41(4): 42-47.
-
文章历史
- 收稿日期:2005-01-04
-
作者相关文章


2. 洛阳市周山森林公园 洛阳 471000;
3. 洛阳市退耕还林工程管理中心 洛阳 471000
2. Zhoushan Forest Garden of Luoyang Luoyang 471000;
3. Management Center of Conversion of Farmland to Forests Project in Luoyang Luoyang 471000
泡桐是我国平原农区与农作物间作的重要速生用材树种,也是出口创汇的主要树种,然而在生产中泡桐的“冠大干低”问题,不仅直接影响到桐材的产量和质量,而且也影响到农作物的产量和品质。为此,从20世纪60年代开始,人们就通过人工接干等方法探讨了泡桐的高干材培育技术(河南农学院泡桐研究室等,1982;1983;蒋建平等,1988;蒋建平,1990;侯元凯等,1999a;范国强等,2000);调查了四川资中地区不同泡桐种类产地的越冬差异(竺肇华,1981);尝试了土平埋保护顶芽越冬的方法(张存义等,1981;侯元凯等,2000);研究了泡桐顶芽的耐寒性和水分变化(侯元凯等,2001;2002)以及泡桐树冠结构、生长性状等对主干以及干形改良的影响(蒋建平等,1988;赵丹宁等,1995a;1995b;崔永兰等,2001)。多数认为泡桐顶芽不能越冬才形成假二叉分枝导致“冠大干低”的,也有认为在寒害到来时期泡桐尚未封顶或未完全木质化造成顶芽枯死的(蒋建平等,1990;侯元凯等,1999)。刘震等(2004)在研究了泡桐顶侧芽休眠发育的温度特性后得出,泡桐顶芽枯死可能是进化过程中采取的一种适应策略,而不是由于冬季寒冷冻死的结果,通过保护顶芽越冬来实现泡桐的自然接干是不现实的,利用泡桐侧芽进行接干应是进行人工接干的主要方法。休眠侧芽萌发形成枝条叫先发枝(proleptic branches),随着主干生长腋芽同时萌发形成枝条叫同发枝或后发枝(sylleptic branches),二者均受到遗传和环境互作的影响,都对树冠结构产生重要影响(原襄,1994;邬荣领等,2002)。因为芽的休眠与萌发成枝反映了树木生长发育的基本单元,树冠与树干的形成都是靠芽的萌发来完成的,并且不同树种即使同一树种不同生态类型在不同气候条件或立地条件下产生不同的反映,表现不同的冬休眠与萌发特性(刘震等,1998a;1998b;2004;刘震,2000;永田洋等,1994;Vegis,1964;Villiers,1975)。为了有效地利用侧芽进行人工接干,人为控制树冠结构,解决泡桐“冠大干低”问题,首先需要对泡桐侧芽萌发成枝接干规律有个清醒的认识。尽管有过兰考泡桐(Paulownia elongata)不定芽接干规律的调查(侯元凯等,1999b)和利用侧芽进行人工接干技术的研究(蒋建平,1990),然而至今未见有关泡桐侧芽萌发成枝规律方面的报道,使得人工接干技术缺乏理论指导。为此,选择生产中推广的杂交种毛白33泡桐(Paulownia tomentosa ×P. fortunei 33)作为研究对象,进行了泡桐侧芽萌发成枝接干规律的研究。
1 材料与方法 1.1 试验地概况试验地位于郑州市须水苗圃(34°43′N,113°39′E)。暖温带大陆性气候,四季分明,年平均气温14.2 ℃,极端最高温度43 ℃,极端最低温度-17.9 ℃,≥10 ℃积温4 717 ℃,无霜期215 d,年平均降水量650.1 mm,全年日照时数大约2 400 h。土壤为潮土类的砂土,微碱性,pH值8.0,有机质含量7.6 g·kg-1, 全氮含量0.58 g·kg -1, 全磷含量1.34 g·kg-1, 全钾含量22.8 g·kg-1。
1.2 试验材料与设计在2000年春季用毛白33泡桐新品种进行埋根育苗,株行距为1.0 m×0.5 m,面积为0.3 hm2,然后任其自然生长,不进行人工修枝、施肥等活动。于2002年4月,按照1%的抽样率,选取泡桐林中间相隔8行的2个南北标准行的50个单株,调查1年生苗干上侧芽在第2年的萌发成枝规律。
1.3 调查内容与方法对每一单株分别调查其主干高Hz、枯梢长度Ls、接干高Hj、胸径D1.3、接干单株的接干处主干直径Dz、接干直径Dj、接干状况(如图 1所示,分通直接干ST、部分错位PT、完全错位FT 3种)、侧芽萌发成枝(包括枯死枝)的长度、粗度、与主干上方夹角等。同时,调查了2001年种植的1年生毛白33泡桐苗(株行距4.0 m×5.0 m)的分枝特性以及接干单株在3~4年后接干通直圆满程度。

|
图 1 泡桐接干状况分类图 Fig. 1 Classification of trunk extension in paulownia trees |
因为泡桐顶芽枯死,由顶芽下第3或第4对侧芽萌发成枝或接干,从而形成泡桐树冠和树干。过去一般认为泡桐分枝为假二叉分枝,这里根据泡桐分枝的实际情况,重新把泡桐分枝类型划分为3种:假二叉分枝、合轴分枝、假单轴分枝(见图 2)。上部一对成活侧芽萌发成枝, 2侧枝与主干夹角大于10°, 表现为二叉状分枝,但泡桐二叉分枝是因顶芽枯死导致的,故命名为假二叉分枝,这也是大家公认的泡桐分枝特性。然而,上部对生成活侧芽由于各种原因导致不能全部萌发,或萌发后2对生侧枝竞争结果导致1侧枝长势过弱或死亡,另1个侧枝进行接干使得泡桐树干向上生长,与互生叶序树木的合轴分枝或具有顶芽接干优势的单轴分枝相似,再把上述泡桐分枝特性判断为假二叉分枝显然不妥。为此,把顶端成活侧芽萌发成枝接干与主干夹角在5°~10°范围内的分枝定为合轴分枝,而把顶端成活侧芽萌发成枝接干与主干夹角为0°的分枝定为假单轴分枝。

|
图 2 泡桐分枝类型 Fig. 2 Types of branching in paulownia trees |
图 3是3种分枝类型所占的比率。由图 3可知,1年生毛白33泡桐最上成活侧芽萌发成枝表现为假单轴分枝的比例最大, 达到44%,合轴分枝最小,仅为14%,二者合计比率达到58%,而假二叉分枝所占比率为42%。同时,在4 m×5 m株行距下毛白33泡桐1年生苗分枝特性调查结果显示,其假单轴分枝与合轴分枝的株数仅占13%。假单轴分枝由于接干枝没有对生侧芽萌发的弱枝竞争,接干枝与主干夹角为0°,可以实现通直接干;合轴分枝表现为对生侧芽萌发枝条发育一强一弱,强枝与主干夹角较小,虽然不能象假单轴分枝的单株那样实现通直接干,但也可以进行较通直的接干;假二叉分枝由于存在着对生侧芽萌发枝条间的竞争,而每一枝条都不能实现向上直立生长,不能实现通直自然接干。所以,在高密度条件下有58%的泡桐单株可以实现自然接干,而在低密度条件下能够实现接干的株数仅占13%。说明高密度可以提高1年生毛白33泡桐最上成活侧芽萌发成枝表现为假单轴分枝和合轴分枝的比例,从而促进泡桐自然接干。尽管如此,即使高密度条件下假二叉分枝的单株比率也达到42%。说明通过增加桐林密度可以提高自然接干单株所占的比率,但不能从根本上解决泡桐由于假二叉分枝导致不能接干或接干不良的问题。

|
图 3 1年生毛白33泡桐苗干上最上成活侧芽萌发成枝的分枝类型及其所占比例 Fig. 3 The proportions of different types of branching sprouted from top lateral buds on trunks of 1-year-old seedlings in Paulownia tomentosa×P.fortunei 33 |
假单轴分枝与合轴分枝的单株平均枯梢长度7.6 cm,而假二叉分枝单株的平均枯梢长度为11.1 cm,对二平均值进行差异显著性检验得出,t=1.429 9<t0.05=2. 045, 说明上侧芽萌发成枝的位置对接干具有一定的影响,但不显著。
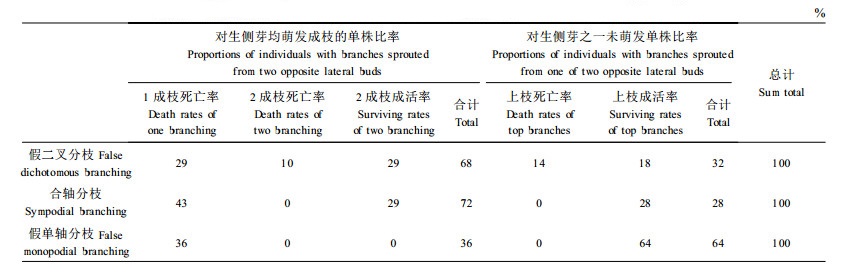
表 1反映了不同分枝类型上侧芽萌发成枝差异。从表 1可见,在假二叉分枝的单株中,有68%的单株树干上部对生侧芽萌发成枝,其中2侧枝均成活的单株占29%,死亡1侧枝的单株占29%,2侧枝都死亡的占10%。但是仍有32%的单株树干上部对生侧芽中只有1个侧芽萌发成枝的,其中侧枝死亡的单株占14%,成活的占18%。上述结果说明不能很好接干的假二叉分枝主要是由2对生侧芽萌发成枝引起的,占到2/3,另1/3单株树干上部对生侧芽中只有1个侧芽萌发成枝,但与主干夹角仍然偏大,无法实现自然接干;在合轴分枝的单株中,有72%的单株树干上部对生侧芽萌发成枝,其中2侧枝均成活的单株占29%,死亡1侧枝的单株占43%。但是仍有28%的单株树干上部对生侧芽中只有1个侧芽萌发成枝,并且成活;在假单轴分枝单株中,树干顶部成活2对生侧芽萌发成枝的单株与1对生侧芽萌发成枝的单株分别占36%、64%,并且全部2对生侧芽萌发成枝单株的1个侧枝死亡,另1侧枝成活而接干。从上述3种分枝类型的顶端成活侧芽萌发成枝差异中可以得出,假二叉分枝与合轴分枝的单株中有2/3以上是2对生侧芽都萌发成枝,只有1/3以下的单株2对生侧芽中仅1个侧芽萌发成枝,而假单轴分枝却相反,2/3单株2对生侧芽中仅1个侧芽萌发成枝,1/3单株2对生侧芽都萌发成枝,并且死亡1个侧枝后仅剩1个侧枝接干,说明要实现泡桐的自然接干需要抑制对生侧芽中1个侧芽的萌发才可能实现,而且也不能保证100%都能接干,因为在树干顶端对生侧芽中即使只有1个侧芽萌发,也存在着分枝夹角大于10°的单株,这可能是单株在群体中与别的单株竞争的结果。
|
|
对假单轴分枝与合轴分枝的接干状况进行分析,得出如图 4、5所示的结果。从图 4看出,接干高与主干高的比(Hj/Hz)在0.3~0.6之间的分布频率较高,达到80%,也就是说80%的接干单株中的接干高在主干高的30%~60%范围内变动;由于泡桐接干的特殊性,在图 5中的接干形率(Dj/Dz)采用接干基径与接干处主干直径的比来表示泡桐接干处的圆满度。从图 5可以看出,接干形率分布在0.8~1.0之间的频率接近达到80%,在0.7~0.8之间有17.2%的分布,在0.6~0.7以及1.1~1.2范围内也有少量分布。说明泡桐侧芽进行自然接干状况下,接干形率较大,可以实现圆满接干,同时有少量单株接干形率较小或过大。

|
图 4 接干高与主干高比的频率分布 Fig. 4 Frequency distribution of ratios of extension height to main trunks height |

|
图 5 泡桐接干形率的频率分布 Fig. 5 Frequency distribution of form quotient of trunk extension in paulownia trees |
图 6显示了毛白33泡桐在第2年接干单株中的接干效果。干形通直,接干效果良好,实现通直接干的占14%;部分错位,接干较差的占28%;完全错位,接干效果最差的占58%。说明第2年侧芽接干能够完全实现通直接干的单株较少,大多数单株即使接干也存在错位的现象。根据对3~4年生接干状况调查的数据,接干的错位状况需要2~3年后才能实现通直圆满。

|
图 6 1年生毛白33泡桐上侧芽萌发成枝接干状况 Fig. 6 Ratios of 3 types of trunk extension sprouted from top lateral buds on trunk of 1-year-old seedlings in Paulownia tomentosa×P.fortunei 33 |
综合上述结果可以得出,在高密度条件下,毛白33泡桐能够靠侧芽接干的单株可以达到58%的高比率,接干形率较大,多数在0.8~1的范围内,然而由于泡桐接干为侧芽接干,第2年能通直接干的比率只有14%,86%的接干存在错位状况。
2.2 下侧芽萌发成枝规律对顶端第一对成活侧芽萌发成枝接干单株的下侧芽萌发成枝的长度、粗度和各侧芽位置成枝数进行分析, 得出如图 7所示的结果。从图 7可见,下侧芽萌发成枝在长度、粗度、成枝数方面变化趋势相同,从上到下均为减小趋势;第1~3对侧芽萌发成枝的长度、粗度、成枝数在个体间变动幅度较大,第4对侧芽以下个体间变动幅度逐渐变小。

|
图 7 2年生毛白33泡桐接干单株中接干枝以下的下侧芽萌发枝条的长度、基径和侧枝的数量 Fig. 7 Length, base diameter and number of branching sprouted from lower lateral buds of 2-year-old seedlings which had extended their trunks in Paulownia tomentosa×P.fortunei 33 |
图 8为1年生毛白33泡桐顶端第1对成活侧芽萌发成枝接干单株的下侧芽萌发成枝与主干上方向的夹角。可以看出,下侧芽萌发成枝夹角平均值自上而下先增后减,呈抛物线型,变化幅度在55°~69°范围内,第4对侧枝与主干夹角最大,达69°;从上到下各侧芽成枝夹角个体间的变动幅度越来越小,分布在较上的侧芽萌发成枝夹角个体间变动幅度较大,最大在30° ~90°范围内,而第6~7对侧芽萌发成枝夹角个体间几乎没有变化。

|
图 8 2年生毛白33泡桐接干单株中接干枝以下的下侧芽萌发枝条与主干上方向的夹角 Fig. 8 Angles up to trunk of branching sprouted from lower lateral buds of 2-year-old seedlings which had extended trunks in Paulownia tomentosa×P.fortunei 33 |
图 9表示下侧芽萌发成枝后的枯死率。可以看出,从上向下,下侧芽萌发成枝的枯死率逐渐增加,第4对侧芽萌发的枝条枯死率已经达到96%,第5~7对侧芽萌发的枝条全部死亡。尽管如此,第1对侧芽萌发形成枝条的枯死率也达到了64%。该结果说明了在高密度种植条件下,泡桐自然整枝强烈。

|
图 9 2年生毛白33泡桐树干下侧芽萌发成枝的枯死率 Fig. 9 The death rates of branches sprouted from lower lateral buds on trunk of 2-year-old seedlings in Paulownia tomentosa×P.fortunei 33 |
综合图 7~9的结果,第4对下侧芽处是下侧芽萌发成枝长度、粗度、侧枝数和夹角变化趋势的共同转折点,该转折点正好说明了泡桐个体不同部位侧芽萌发成枝后生长发育的差异。为争夺光照资源第1~3对下侧芽萌发后向不同方向生长成枝,个体间差异明显,而越往下侧枝受到上部侧枝遮荫越严重,为了争夺光照资源,只有靠增加与树干夹角进行横向发展,到第4对侧枝夹角达到最大,结果第4对以上的侧芽可以得到充足的阳光,生长发育良好。然而再往下的侧枝由于受到第4对以上侧枝的遮荫,生长受到明显的抑制,表现为枝条长度、粗度、侧枝数均降低,与树干上方向夹角也逐渐变小。上述结果说明上部侧芽具有明显的萌发成枝优势,对第4对以下侧芽萌发成枝具有一定的抑制作用。
3 讨论1年生毛白33泡桐侧芽在第2年春季萌发成枝接干与成冠。由于芽的发育或竞争,使上部对生侧芽同时萌发形成均势的假二叉分枝;或者发育成一强一弱,与主干夹角小于10°的合轴分枝;或者弱枝枯死,强枝接干,或者仅萌发一个侧芽成枝接干,接干枝与主干夹角为0°的假单轴分枝。把泡桐的分枝类型划分为这3种类型更切合实际,因为泡桐的分枝类型本来就不是假二叉分枝1种,如果把这3种类型归为1类的话,也只能归为假轴分枝(原襄,1994),因为这3种分枝都是侧芽萌发后,或向主轴靠拢形成合轴分枝与假单轴分枝,或向斜上方生长成二叉分枝,都没有顶芽萌发形成的主轴,只能算是假轴,假二叉分枝只是假轴分枝的1种而已。
在高密度情况下,毛白33泡桐分枝习性表现为合轴分枝与假单轴分枝,实现自然接干的比例高达58%,而在低密度条件下能够实现自然接干单株的比率仅13%,说明高密度下个体间竞争加剧,利于泡桐接干,但高密度不能改变泡桐牺牲顶芽尽量延长生育期的生态适应特性(刘震等,2004),使具有假轴分枝特性的泡桐单株都能进行假单轴或合轴分枝,实现自然接干。同样,分布于亚热带气候自然接干表现较好的白花泡桐能够实现自然接干的比率较高只是合轴分枝与假单轴分枝比率较高而已。采用人工接干是解决泡桐“冠大干低”培育高干桐材的主要途径(刘震等,2004)。
实现自然接干的泡桐单株多是在顶芽下第3~4对越冬对生休眠侧芽(刘震等,2004)中只有1个萌发成枝接干,另一个侧芽要么不萌发,要么萌发后长势弱,甚至死亡。自然状态下泡桐接干形率虽然较高,但接干高度还多在主干高的30%~60%,并且第2年能通直接干的比率也只有14%,86%的接干存在错位状况,且要在2~3年后才得以改善,才能像利用顶芽接干的单轴分枝杨树那样实现通直圆满。
接干侧芽下第4对侧芽是上下侧芽萌发成枝生长的转折点,第4对侧芽以上萌发枝条的长度、粗度都较大,而以下侧枝则较小,自然整枝强烈,并且第4对侧枝与主干夹角达到最大。所以,进行抹芽接干时最好抹除第4对以下侧芽,以防止养分的无效消耗。
崔永兰, 吕国政, 石俊阁. 2001. 泡桐接干类型划分及干形相关性状的分析. 河南农业大学学报, 35(1): 70-73. DOI:10.3969/j.issn.1000-2340.2001.01.020 |
范国强, 王安亭, 王国周, 等. 2000. 接干和施肥对不同初植苗高泡桐幼树主干生长影响的研究. 林业科学研究, 13(6): 628-633. DOI:10.3321/j.issn:1001-1498.2000.06.009 |
河南农学院泡桐研究室, 禹县林业局, 禹县林科所. 1982. 泡桐剪梢接干效果的分析. 河南农学院学报, 16(4): 1-9. |
河南农学院泡桐研究室, 禹县林业局, 禹县林科所. 1983. 泡桐剪梢接干法的技术要点. 河南农学院学报, 17(1): 27-28. |
侯元凯, 翟明普. 1999a. 泡桐干形培育研究. 林业科学, 35(3): 76-83. |
侯元凯, 翟明普, 娄季松, 等. 1999b. 兰考泡桐不定芽自然接干规律研究. 北京林业大学学报, 21(3): 14-19. |
侯元凯, 翟明普, 张俊昌, 等. 2000. 兰考泡桐苗木顶芽越冬特性初步研究. 河南农业大学学报, 34(2): 196-200. DOI:10.3969/j.issn.1000-2340.2000.02.028 |
侯元凯, 翟明普, 聂爱社, 等. 2001. 兰考泡桐苗木顶芽水分变化规律研究. 北京林业大学学报, 23(6): 17-21. DOI:10.3321/j.issn:1000-1522.2001.06.005 |
侯元凯, 翟明普, 聂爱社, 等. 2002. 兰考泡桐苗木顶芽耐寒性研究. 中国生态农业学报, 10(2): 19-21. |
蒋建平, 李荣幸, 刘廷志. 1988. 泡桐树冠结构对主干生长影响的研究. 河南农业大学学报, 22(1): 1-9. |
蒋建平主编.1990.泡桐栽培学.北京:中国林业出版社, 263-283
|
刘震, 何松林, 王艳梅, 等. 2004. 泡桐顶侧芽休眠发育的温度特性研究. 林业科学, 40(3): 46-50. |
刘震, 王玲. 2000. 不同种源山桐子冬芽休眠的温度特性. 河南农业大学学报, 34(3): 252-254, 297. DOI:10.3969/j.issn.1000-2340.2000.03.015 |
邬荣领, 胡建军, 韩一凡, 等. 2002. 表型可塑性对木本植物树冠结构与发育的影响. 林业科学, 38(4): 141-156. DOI:10.3321/j.issn:1001-7488.2002.04.023 |
张存义, 殷年喜. 1981. 泡桐新的接干办法--顶芽接干. 安阳林业科技, (1): 2-3. |
赵丹宁, 熊耀国, 宋露露. 1995a. 泡桐树冠结构与生长性状遗传相关的研究. 西北林学院学报, 10(4): 11-16. |
赵丹宁, 熊耀国, 宋露露, 等. 1995b. 白花泡桐树冠结构、生长性状的选择对干形改良的影响. 林业科学研究, 8(1): 82-87. |
竺肇华. 1981. 泡桐属植物的分布中心及区系成分探讨. 林业科学, 17(3): 271-279. |
刘震, 栉田达矢, 永岛敦司, 等. 1998a. イイギリの成长停止を诱导する环境条件に关する研究. Environmental Engineering Research, 35: 487-494. |
刘震, 永岛敦司, 栉田达矢, 等. 1998b. 冬季の温度影响がイイギリの开芽に及ぼす影响. Environmental System Research, 26: 239-244. |
刘震. 2000. 亚热带域に分布するイイギリの休眠に关する研究. 三重大学演习林报告, 24: 107-161. |
永田洋, 中岛敦, 万木丰. 1994. 树木の芽の休眠. 三重大学演习林报告, 18: 17-42. |
原襄. 1994. 植物形态学. 初版.东京: 朝仓书店, 13-35.
|
Vegis A. 1964. Dormancy in higher plants. Annual Review of Plant Physiology, 15: 185-224. DOI:10.1146/annurev.pp.15.060164.001153 |
Villiers T A. 1975. Dormancy and the survival of plants. London: Edward Arnold.
|