 2005, Vol. 41
2005, Vol. 41文章信息
- 苏瑞军, 苏智先.
- Su Ruijun, Su Zhixian.
- 珙桐种子散布、萌发及其种群龄级分配的关系研究
- Study on Relations of Seed Dispersal, Germination and Age-Class Distribution of Population in Davidia involucrata
- 林业科学, 2005, 41(3): 192-195.
- Scientia Silvae Sinicae, 2005, 41(3): 192-195.
-
文章历史
- 收稿日期:2004-01-12
-
作者相关文章


2. 绵阳师范学院 绵阳 621000
2. Mianyang Normal College Mianyang 621000
珙桐(Davidia involucrata)为我国单属种的特有植物。珙桐起源古老, 系第三纪古热带植物区系的孑遗种, 在植物系统发育和地史变迁研究上有很高的学术地位, 被列入国家一级保护珍稀濒危植物。此外, 珙桐开花时其头状花序基部有2枚白色的大苞片, 随风飘动, 如白鸽展翅, 有“中国鸽子树”之称, 有极高的观赏价值, 是重要的园林绿化树种(贺金生等, 1995; 傅立国等, 1992)。过去对珙桐的研究涉及到许多方面, 如它的分类地位(Li, 1954)、引种繁殖(唐晓军, 2002; 沈作奎等, 1998; 张家勋等, 1995)、解剖特征(孙彬等, 1993)、生殖生物学(苏智先等, 1999)、化学成分(向桂琼等, 1989)、群落学特性(杨业勤等, 1986; 钟章成等, 1984; 杨一川等, 1989; 李傅等, 1990)等方面; 但对珙桐种子在自然状况下的散布、萌发和其种群龄级配备关系的研究很少。加上珙桐的种子种壳厚而坚硬且后熟期长, 一般要2~3年才能破壳发芽, 大多数在发芽前就已腐烂, 并且有鼠类等动物啃食种子。所以研究自然条件下它的种子散布、萌发及种群配备的关系对珙桐种群更新的研究及其群落的有效保护具有重要的意义。卧龙自然保护区三江镇鹿尔坪村的白泥岗生长着世界最大的珙桐群落。本文是对这里所分布的珙桐种群进行野外调查结果的报道。
1 研究地点自然概况研究地点在卧龙自然保护区三江镇鹿尔坪村的白泥岗, 卧龙地区海拔在1 150~6 250 m。卧龙自然保护区位于成都平原到西藏高原的过渡斜面上, 阿坝藏族自治州部汶川县境, 岷江上游, 邛崃山脉之东南部, 属于高山峡谷地段。本区属青藏高原气候区的范围, 西风急流南支和东南季风控制本区的主要天气过程。该地区气候凉爽, 干湿季节明显。冬半年天气晴朗干燥, 降雨量少; 夏半年东南季风经岷江河谷溯流而上, 受邛崃山脉迎风坡的阻挡, 雨量丰沛, 气候凉爽。根据沙湾(海拔1 920 m)气象观测资料, 卧龙年均温8.4 ℃, 1月均温-1.7 ℃, 7月均温17.0 ℃。年均降水量861.10 mm, 5—9月降水量占全年的68.1%, 年平均相对湿度80%, 年日照时数926.7 h, 土壤为山地黄棕壤, pH在5.8~6.5, 原生林内枯枝落叶层达3~6 cm(魏东峰, 1997)。
白泥岗样地位于102°56′E, 30°51′N的一座山的南坡, 珙桐群落主要分布在海拔1 450~2 200 m。由于人类活动如森林砍伐、放牧、采药等的影响, 使珙桐群落在距居住地较近的地方遭到了严重的破坏。
2 研究方法调查时间为2003年4—6月。利用群落样方法(王伯荪等, 1996)在珙桐群落中心设置横向8个连续的样地和纵向8个连续的样地。乔木样地沿水平方向设置, 大小为20 m×20 m。灌木样方设置于乔木样地内, 调查4个大小为5 m×5 m的样方。样地内各林木的年龄是通过调查前和当地农民及保护区的工作人员的座谈来确定的。在16个样地中找到12棵开花的大树, 以树基部为中心, 以大于冠幅1 m为半径做圆形样方。在坡度大的地方, 据实际情况适度的将坡上向上缩短和将坡下向下延伸1 m。先收集样方内草本层中裸露的种子, 然后在腐质层10 cm深处, 收集所有的种子。对种子的完整性及幼苗和幼树进行统计。
调查时记录样方的面积、海拔、坡度、坡向、总盖度(包括乔、灌、草分层盖度)干扰等情况。对乔木进行每木检尺。调查其高度、胸径、冠幅、枝下高、坐标, 数量。
3 结果与分析 3.1 珙桐种子的散布行为珙桐的果实为核果, 呈长椭圆形, 径3~5 cm, 具肉质果皮, 内果皮骨质, 表面有深沟, 十分坚硬, 内含1~6粒种子。种子较大, 长1.5~2.5 cm, 宽1.5~1.7 cm。果实大而呈椭圆形使其在成熟落地时易滚动, 其分布因母树所处的地势而定。一般, 在树根部较多, 若地势较陡, 果实落地后可能由于雨水等作用而聚集在树下方的沟崖处。其分布范围内种子的密度很大程度上取决于地势。一般在离树基部5 m较集中, 若地势较陡则离树冠垂直较远。种子的高度密集和落地随地势的滚动大大限制了种群的自然扩张。种子刚落地时是裸露的, 由于珙桐林的土壤属坡积土、结构松散, 再加上雨水的冲刷和人为及家畜动物的活动, 造成部分种子埋到土层中, 但深浅不一, 总的约有7%的种子裸露在地表。调查结果见图 1, 1株珙桐树散布的所有种子的80%~90%集聚在离母树10 m的范围内, 几乎都是在母树的树冠之下。

|
图 1 珙桐种子散布图 Fig. 1 Seed dispersal range of Davidia involucrata |
在调查的过程中发现, 松鼠等动物对珙桐种子的散布也有一定的影响。在石头下发现的2堆松鼠贮藏的果核, 其中野核桃的果核占65%, 珙桐的占35%, 由于其贮藏的环境比较干燥, 珙桐种子在这种情况下也是很难萌发的。
3.2 种子的萌发情况种子是珙桐种群得以形成的2种因素之一。珙桐种子的萌发从形态结构和种子生理学角度看都是很迟才萌发。一般成熟时从树上掉落到地面要经过2年左右才能完成种子的后熟, 从而才有机会突破内果皮而萌发。珙桐的自行繁殖有很大的困难。在落地后的2~3年内, 种子在雨水的冲击下, 有的被埋到了地下, 造成了85%以上的种子腐烂, 再加上虫子的危害, 能发芽的很少。
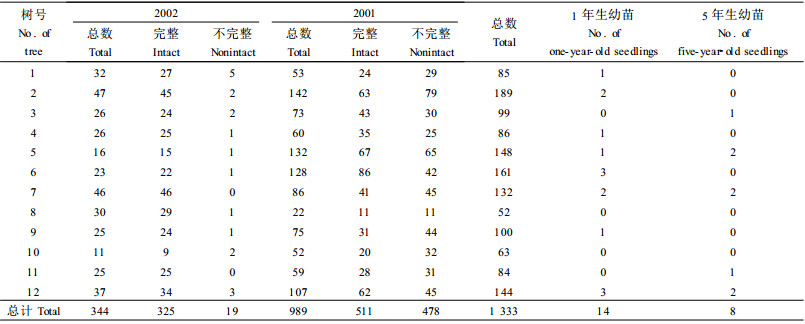
从表 1看出, 在每株2002年成熟的种子经过1年时间的变化, 由于有内果皮及外果皮的保护, 不完整的占全部种子的1%~36%;每株2002年以前的不完整的种子占全部种子的21% ~53%。有一半左右的种子不完整。1年生幼苗是由2001的种子萌发而来的, 萌发的种子仅占全部种子的0~2.1%。每年仅有最多2.1%的种子萌发出幼苗, 种子萌发后进入漫长的营养生长期。其中, 大量的幼苗由于过湿或其他不利的环境胁迫, 导致幼苗大量的死亡, 并没有生长发育到幼树和成长起来。在一个多月的调查中, 沿途碰到的幼苗有8株, 刚长出的2片子叶很脆易折, 其中被家畜、野生动物或挖药的农民踩倒踩死的有3株。过度的人为和家畜活动对珙桐幼苗的成活影响也很大。
|
|
珙桐种子的散布方式对其幼苗的分布起着决定性作用。如前所述, 种子主要散布在母树的附近, 其发育成长起来的幼树也主要生长在树的周围。在调查中的14棵幼苗和8棵幼树, 除了4株幼树外, 其余的都分布在大树的周围, 其生态幅狭窄, 生长受剑种群竞争的压力很大。受着上述的各种死亡因素的影响, 靠种子更新和基部砍伐萌生的珙桐种群扩张有很大的局限性。
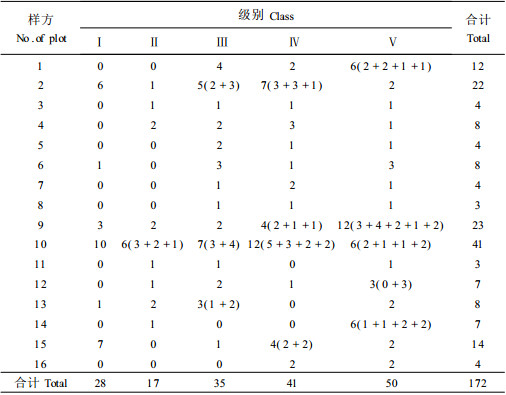
3.3 珙桐径级结构对12个样地的珙桐树以径级结构代替年龄结构分析。分级标准:Ⅰ级幼苗高H < 0.33 m; Ⅱ级幼树H>0.33 m, 胸径DBH < 2.5 cm; Ⅲ级小树DBH2.5~7.5 cm; Ⅳ级中树DBH7.5~22.5 cm; Ⅴ级大树DBH>22.5 cm。表 2各样地中珙桐分级株数的统计, 其中6(2+2+1+1)表示6株里面有4株是2株1丛的, 有2株是单株的。
|
|
由于珙桐为国家一级保护树种, 所以不能轻易用生长锥法或解析树干法测其实际年龄。在外形上也很难观测出其年龄。因此在调查中我们采用实际测定各个珙桐个体的胸径, 用径级百分比近似代替珙桐的年龄结构(艾训儒等, 1999)。
图 2可以看出, 各样地中各级的株数都不一样。在总的样地中Ⅴ级的大树最多, 占到了29%, Ⅳ级的中树占24%, Ⅲ级的小树占20.2%, Ⅱ级幼树的最少, 仅占9.8%。Ⅰ级的幼苗占16%。其中大中树占到了一半以上, 幼树短缺, 不过幼苗有所回升。但从表 2可以看出, 样地中幼苗主要出现在3个样地内:样地2中有6株, 样地10中有10株, 样地15中有7株, 共23株, 占总数的82.1%。总体上各级从大小排列, 呈倒金字塔排列, 珙桐种群中幼树、幼苗的缺乏使得珙桐种群处于衰退。

|
图 2 珙桐径级结构 Fig. 2 The DBH structure of Davidia involucrata population |
对实地数株珙桐年轮的观测, 以及参考有关文献资料, 珙桐的胸径与其年龄之间存在着一定的相关关系。根据这种关系可将图 2中的数据换算为表 3的年龄数。
|
|

由表 3得知, 从种子到幼苗的成苗率只有1.01%, 说明其萌发困难及萌发后的营养生长过程中死亡率较高。而从其他各龄级的保存率及它们所占种子的百分率看, 都大于50%, 甚至有的达到200%。说明靠种子萌发而长成的大树很少, 多数是靠砍伐后基部萌蘖而长成的。由于农民非法采集种子, 使珙桐种子库里面的数量减少; 另一方面就是在山上挖取幼苗。再加上以前大量的砍伐使的珙桐原始种群在不断减少, 出现了幼苗幼树缺乏的现象, 种群的更新困难。这样使得珙桐种群整体处于衰退状态。
4 讨论本文认为, 目前的珙桐种群龄级分配不很理想。种群靠自生种子和萌蘖繁殖的自我更新缓慢, 更新能力较弱, 更新范围局限在树体1~10 m环带附近, 使种群难以扩张。再加上珙桐种子保存率和幼苗成活率都较低, 这些状况使珙桐种群处于濒危。因此, 必须加强对珙桐种群的自然保护, 防止人为严重干扰, 禁止砍伐。同时研究在人工控制条件下珙桐种子的生物学特性以及人工林培育的科学管理方法, 弄清珙桐种群濒危的原因并作出相应的拯救措施, 以便珙桐这一稀有珍贵的资源得以较好地保存。
艾训儒, 谭建锡. 1999. 星斗山自然保护区珙桐种群结构特征研究. 湖北民族学院学报(自然科学版), 17(1): 12-15. |
傅立国, 金鉴明. 1992. 中国植物红皮书——稀有濒危植物(第一册). 北京: 科学出版社, 474-475.
|
贺金生, 林洁, 陈伟烈. 1995. 我国珍稀特有植物珙桐的现状及其保护. 生物多样性, 3(4): 213-221. DOI:10.3321/j.issn:1005-0094.1995.04.005 |
李傅, 袁道凌. 1990. 鄂西七姊妹山珙桐群落及其保护对策研究. 华中师范大学学报(自然科学版), 24(3): 221-323. |
沈作奎, 艾训儒. 1998. 星斗山自然保护区珙桐繁殖方式及生长分析. 湖北林业科技, (4): 1-3. |
孙彬, 李柏年, 林障德, 等. 1993. 两种珙桐叶片结构的观察. 西北植物学报, 13(3): 198-202. DOI:10.3321/j.issn:1000-4025.1993.03.006 |
苏智先, 张素兰. 1999. 珙桐种群生殖物候及其影响因子研究. 四川师范学院学报(自然科学版), 20(4): 313-318. |
唐晓军. 2002. 珙桐的引种繁殖技术. 林业科技开发, 16(3): 51-53. DOI:10.3969/j.issn.1000-8101.2002.03.025 |
王伯荪, 余世孝, 彭少麟, 等. 1996. 植物群落学实验手册. 广州: 广东高等教育出版社.
|
魏东峰. 1997. 卧龙自然保护区大气环境现状分析与评价. 四川林业科技, 18(3): 75. |
向桂琼, 卢馥菘. 1989. 中国特有植物珙桐不同化学成分研究. 植物学报, 31(7): 540-543. |
杨业勤, 徐友源. 1986. 贵州珙桐生态特性的初步研究. 林业科学, 22(4): 426-430. |
杨一川, 李体俊. 1989. 四川峨眉山的珙桐群落的初步研究. 植物生态学与地植物学丛刊, 13(3): 270-276. DOI:10.3321/j.issn:1005-264X.1989.03.001 |
张家勋, 李俊清. 1995. 珙桐繁殖和栽培技术研究. 北京林业大学学报, 17(3): 24-29. |
钟章成, 秦自生, 史建慧. 1984. 四川卧龙地区珙桐群落特征的初步研究. 植物生态学与地植物学丛刊, 8(4): 253-263. |
Li Huilin. 1954. Davidia as the type of a new family. Dvidiaceae Lloydia, 17(4): 329-331. |