 2005, Vol. 41
2005, Vol. 41文章信息
- 谢钰容, 周志春, 廖国华, 金国庆, 陈跃.
- Xie Yurong, Zhou Zhichun, Liao Guohua, Jin Guoqing, Chen Yue.
- 低磷胁迫下马尾松种源酸性磷酸酶活性差异
- Difference of Induced Acid Phosphate Activity Under Low Phosphorus Stress of Pinus massoniana Provenances
- 林业科学, 2005, 41(3): 58-62.
- Scientia Silvae Sinicae, 2005, 41(3): 58-62.
-
文章历史
- 收稿日期:2003-05-12
-
作者相关文章



2. 福建省三明市林业局 三明 365000;
3. 福建省南纸股份有限公司 南平 353000
2. Sanming Forest Bureau of Fujian Province Sanming 365000;
3. Fujian Nanping Paper Co.Ltd. Nanping 353000
土壤有效磷的缺乏是一种极为普遍的现象。为有效利用低浓度的土壤磷素,小麦、玉米、大豆、黄瓜、番茄等许多作物在形态解剖特征和生理生化特性方面都会发生系列适应性的变化,其中一项重要的变化就是酸性磷酸酶(APase)活性的增强。APase主要存在于植物根的表皮或叶的下表面,或者由根系分泌至外部介质(Lefebvre et al., 1990;Hirata et al., 1982)。APse是一种适应性诱导酶,当植物获得磷饥饿诱导(phosphorus starvation induce,PSl)的信号后, APase活性将显著增强,它能使复杂的有机磷化合物水解为植物易吸收利用的磷酸盐,可使植株体内仅有的磷得到多次重复利用(樊明寿等,2001;丁洪等,1997)。Hirata等(1982)观察到PSI刺激APase分泌到小麦及水稻幼苗的培养液中;Kummerva(1986)发现PSI增加玉米根表面的磷酸酶;Lefebvre等(1990)认为根系APase活性和分泌性APase活性反映植物活化和分解难溶性磷的能力,与植物磷效率关系密切。不同磷营养效率的植物基因型在低磷胁迫下APase活性和分泌性APase活性不同,如吴平等(2001)研究发现耐低磷胁迫水稻品种的分泌性APase活性显著高于磷敏感性品种。研究分析植物APase活性和分泌性APase活性可以从一个层面揭示不同植物基因型对低磷胁迫的适应机制和磷效率的遗传差异。马尾松(Pinus massoniana)不同种源对低磷胁迫的遗传反应差异显著,如福建武平和广东信宜2种源适宜于低磷条件下生长,而广西岑溪等种源对磷肥反应敏感(周志春等,2003)。谢钰容等(2003;2004)报道了马尾松不同种源的磷素吸收动力学参数和根系有机酸分泌物特征,本文则进一步从APase活性和分泌性APase活性角度阐明造成马尾松不同种源对磷肥反应差异的适应机制及与磷效率等关系。
1 材料与方法 1.1 试验材料供试材料为浙江淳安、福建武平、江西崇义、广东信宜和广西岑溪马尾松种源,这些种源分布在亚热带不同地理区域,对磷肥的遗传反应已经研究得比较清楚(周志春等,2003)。盆栽试验基质为中国林业科学研究院亚热带林业研究所虎山的贫瘠缺磷酸性红壤(谢钰容,2003)。
1.2 试验设计 1.2.1 盆栽试验和幼苗水培试验按种源与磷肥2因素完全随机区组设计进行盆栽和水培试验,对应地设置低(缺)磷(P0)、中磷(P1)和高磷(P2)3种磷水平处理,3次重复。具体参见谢钰容(2003)。
1.2.2 根部分泌性APase活性测定采用Mclanchlan(1980)方法测定。2002年7月上旬取水培的1年生幼苗用蒸馏水洗净吸干根系后,置于50 mL预先通气的含有0.5 g·L-1 对硝基苯磷酸二钠(pNPP)的0.2 mol·L-1醋酸钠缓冲液(pH5.0)中。用培养箱室温培养90 min后,立即加入2 mol·L-1NaOH终止酶促反应,于405 nm波长条件下比色测定生成的对硝基苯酚(NPP)的量。剪下幼苗根系于70 ℃烘干至恒重。酶活性以单位时间内单位干质量根水解pNPP生成NPP的量表示,即μg·g-1root DW min-1。同时做空白对照,即在反应液放幼苗前加2 mol·L-1NaOH。
1.2.3 针叶和根系APase活性测定分别于8月13日、9月3日和10月3日利用土培幼苗测定。测定前用蒸馏水把待测定植株洗净吸干,剪下部分针叶和根系称重,并立即加入2 mL冰蒸馏水冰浴研磨成浆,再加入3 mL冰蒸馏水将其全部洗入7 mL离心管中。在4 ℃、9 000 r·min-1条件下离心15 min。取2 mL上清液测定APase活性,测定方法同上。酶活性以单位时间内单位(针叶或根)鲜质量水解pNPP生成NPP的量表示,即μg·g-1root FWmin-1。
1.2.4 光合速率及磷效率测定用CI-310型便携式光合仪在室内人工光源下测定马尾松种源土培苗生长高峰期(8月)光合作用CO2的变化。光合速率以单位时间(s)单位干质量(g)所消耗的CO2量(μg)表示,即μg CO2·g-1DWs-1,试验重复3次。种源的磷效率以其收获时(10月底)低磷与中(高)磷水平下干物质的积累量之比表示,即磷效率=低磷胁迫下干物质积累量/中(高)磷条件下干物质积累量×100%(曹靖等,2000)。
2 结果与分析 2.1 低磷胁迫下马尾松种源分泌性APase活性差异不同磷水平下马尾松种源水培苗根系分泌性APase的活性测定值列于表 1。表 1数据显示,随培养介质磷水平的下降,马尾松种源根系分泌性APase活性显著增加,磷水平间差异达到极显著水平(p<0.000 1),其活性增幅在29%~101%(中磷与高磷比较)和51 %~257%间(低磷与高磷比较),具有明显的趋势性。这意味着低(缺)磷逆境会刺激马尾松种源根系APase向根际的分泌,磷胁迫的程度越严重,其根系APase分泌的能力就越强。
|
|
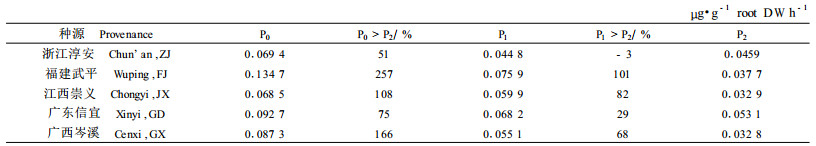
进一步地统计分析表明,不同磷水平下各参试种源根系分泌性APase的活性差异也非常显著(p<0.001 6)。在低磷水平下,福建武平种源分泌性APase活性最强,广东信宜种源次之,而浙江淳安和江西崇义种源最低,其分泌性APase活性仅为福建武平种源的50%左右。在中磷水平下也大致具有同样的变化规律。
2.2 低磷胁迫下马尾松种源针叶和根系APase活性差异低磷胁迫下,马尾松种源除了通过根表分泌APase以活化和分解根际的有机磷供植物吸收利用外,植株体内包括针叶和根系的APase活性也会发生变化,以使植株体内仅有的磷得以多次重复利用。从表 2给出的马尾松生长高峰期(8月)参试种源土培苗根系和针叶的APase活性来看,可清楚地说明APase活性的变化规律,即随着磷胁迫程度的加深,马尾松种源根系和针叶的APase活性将显著提高,表现出其与磷营养水平的负相关性。如低磷水平下,种源根系和针叶APase活性分别较高磷水平提高135%~254%和189%~233%。对照低磷水平下根系和针叶APase的活性,发现前者在总体上都高于后者,这是由于根系是养分进入的门户,较早接受低磷胁迫的信号,对低磷胁迫的敏感性也较强。
|
|

方差分析结果表明,低磷水平下马尾松参试种源的根系APase活性存在显著的遗传差异(p<0.091 6),其中福建武平和广东信宜2种源的根系APase活性最高,较广西岑溪种源高出1倍左右,分别达0.088 7和0.089 8 μg·g-1root FW h-1,而江西崇义、浙江淳安和广西岑溪这3个种源的根系APase活性相对较小,只有0.061 0、0.057 6和0.046 0 μg·g-1 root FW h-1。低磷逆境下土培幼苗针叶APase活性的种源差异类似其根系APase活性变化。这一结果表明,较之于其他3个种源,福建武平和广东信宜2种源在低磷逆境下更能重复利用体内的有机磷供机体新陈代谢之需,以应对低磷胁迫,保证植株的良好生长发育。在中磷和高磷水平下,福建武平和广东信宜种源根系APase的活性也高于其他种源。
2.3 不同生长期马尾松种源根系和针叶APase活性变化本试验还测定了不同生长时期马尾松土培苗根系和针叶APase活性,以研究比较其季节性的变化。图 1结果显示,无论是根系还是针叶,马尾松参试种源在低磷水平下APase活性都表现为8月>9月>10月,即在测定APase的3个时期,8月植株体内APase活性最高,而在生长高峰期过后,马尾松幼苗APase活性急剧下降,到10月下降到最小值,表现出较强的时效性。结合马尾松的季节生长规律,APase活性的这种规律性变化正反映了植株从生长高峰期向休眠期的过渡。在不同生长期,马尾松APase活性的种源差异是不同的。在生长高峰期的8月,种源间APase活性差异极为显著,而至9月和10月,不同种源APase活性未表现出明显的差异。

|
图 1 低磷水平下不同生长期马尾松种源根系(左)和针叶(右)Apase活性 Fig. 1 APase activity in root(left) and leaf (right) of five provenances tested at low phosphorus level during different growth phases(unit of APase activity:μg·g-1 root FW h-1) |
植物APase活性变化反映其体内磷素的重复利用能力,光合作用是植物同化无机物为有机物(包括光合磷酸化和碳循环)能力的体现,而磷效率则指当介质中磷素的有效浓度较低时植物正常生长的能力,即植物耐低磷的能力差异(严小龙等,1992)。对照马尾松种源根系和针叶APase活性与其光合速率和磷效率(表 2)可以看出,在低磷水平下,福建武平种源具有最大的光合速率(8.457 μg CO2·g-1DW s-1)、最强的根系和针叶APa se活性(分别为0.088 7和0.063 0 μg·g-1root FW h-1)、最高的磷效率(P0/P1和P0/P2分别为71.17%和71.74%)和生物量(0.337 2 g)。从低磷胁迫条件下广东信宜种源的根系APase活性、光合速率和磷效率测定值看,该种源的表现也较佳。通过比较表 3中3种磷水平下各参数变化,结合已有研究结果(周志春等,2003;谢钰容等,2003; 2004),可认为福建武平和广东信宜乃2个耐低磷型优良种源,而APase活性、光合速率、磷效率这3个具有较好一致性的指标可用以评价马尾松种源在低磷水平下生长表现。
3 结论与讨论马尾松是我国亚热带地区重要的针叶纸浆原料树种。虽然南方山地土壤有效磷的缺乏是一种普遍存在的现象,但马尾松并不因此表现缺磷而生长良好,这说明其有适应低磷胁迫的应对机制。研究观测到,马尾松根际对土壤中的磷具有活化和富集作用,但不同种源间差异较大,如福建武平和广东信宜2种源对磷肥反应不敏感,活化和富集土壤磷的作用较其他种源强烈(周志春等,1993)。通过水培试验,作者从根系分泌物和磷素吸收动力特征角度已初步阐明了造成马尾松种源适应低磷逆境差异的生理机制(谢钰容等,2003;2004)。
在低磷胁迫下,APase活性和分泌性APase活性的提高是植物适应磷胁迫的重要机制之一。植物接受到磷饥饿的刺激后其根系会向根际大量分泌APase,以水解生长介质中的无效态有机磷释放PO43-(李玉京等,1997)。缺磷逆境还将增强植株APase的活性以提高体内磷的利用率,通过耗用较少的磷产生尽可能多的生物量(徐庆章等,1989)。如Fox等(1992)在研究了湿地松(Pinus elliottii)根际APase与有机磷的转化关系后认为根际APase的增加提高了磷的有效性。基于分根处理,庞欣等(2000)发现,不供磷的黄瓜根系APase活性显著高于供磷根系,该酶活性与根系的含磷量显著相关。
本文结合利用水培试验和土培试验,研究5个对磷肥反应式样不同种源的根际分泌性APase和植株体内APase活性变化,以进一步解释大田试验结果和阐明马尾松种源对磷饥饿的适应机制。水培试验研究结果表明,低磷胁迫将诱导和刺激马尾松种源根系APase的分泌。随着培养介质磷水平的下降,根系分泌性APase的活性显著增加,增幅在29%~101%(中磷与高磷比较)和51%~257%间(低磷与高磷比较)。低磷水平下5个马尾松参试种源根系分泌性APase活性是不同的,以福建武平种源最强,广东信宜和广西岑溪种源其次,浙江淳安和江西崇义种源最低,仅为福建武平种源的50%左右。
土培试验发现,生长高峰期马尾松种源根系和针叶APase活性与磷素水平的关系类似于根系分泌性APase活性变化,即随着介质磷素的减少和磷胁迫的加重,根系和针叶APase的活性越强。同样,与其他参试种源相比,福建武平和广东信宜2种源在低磷胁迫下根系和针叶APase的活性较高,分别达0.088 7、0.063 0和0.089 8、0.045 0 μg·g-1root FW h-1。然而,生长高峰期过后,马尾松根系和针叶APase活性急剧下降,种源差异显著减小。这一结果也意味着,宜选择林木生长的高峰期(7—8月)取样测定,这样才能确切地研究说明不同林木品种和种质在低磷胁迫下的APase活性差异,正确评价其耐低磷的特性。研究还发现,不同磷水平下,尤其是低磷水平下马尾松种源根系APase的活性一般都高于针叶,这说明根系对磷饥饿的敏感性较强。较之于针叶,低磷胁迫下根系APase的活性是一个较好反映林木耐低磷特性的生理指标。
通过比较马尾松不同种源的APase活性、光合作用和磷效率,发现这3者具有较好的一致性,可用来指示和评价马尾松种源的耐低磷特性。根据这3个指标可进一步认定福建武平和广东信宜是2个耐低磷型的优良种源。对于植物的光合速率,缺磷因影响其代谢过程而一般表现为负作用(潘晓华等,1997),但在本试验中观察到这2个耐低磷种源在低磷水平下的光合作用速率高于中磷和高磷水平,这并不意味着低磷水平下2种源就具有最高的干物质积累量。在甘蔗研究中也发现类似的现象(万美亮等,1999),更深层的原因尚待研究。
曹靖, 张福锁. 2000. 低磷条件下不同基因型小麦幼苗对磷的吸收和利用效率及水分的影响. 植物生态学报, 24(6): 731-735. DOI:10.3321/j.issn:1005-264X.2000.06.015 |
丁洪, 李生秀, 郭庆元, 等. 1997. 酸性磷酸酶活性与大豆耐低磷能力的相关研究. 植物营养与肥料学报, 3(2): 123-128. DOI:10.3321/j.issn:1008-505X.1997.02.005 |
樊明寿, 徐冰, 王艳. 2001. 缺磷条件下玉米根系酸性磷酸酶活性的变化. 中国农业科技导报, 3(3): 33-36. DOI:10.3969/j.issn.1008-0864.2001.03.009 |
李玉京, 李滨, 李继云, 等. 1997. 低磷营养胁迫下小麦RNA酶与酸性磷酸酶活性的变化. 西北植物学报, 17(4): 417-425. DOI:10.3321/j.issn:1000-4025.1997.04.001 |
潘晓华, 石庆华, 郭进耀, 等. 1997. 无机磷对植物叶片光合作用的影响及其机理的研究进展. 植物营养与肥料学报, 3(3): 201-207. DOI:10.3321/j.issn:1008-505X.1997.03.002 |
庞欣, 张福锁, 李春俭. 2000. 部分根系供磷对黄瓜根系和幼苗生长及根系酸性磷酸酶活性影响. 植物生理学报, 26(2): 153-158. DOI:10.3321/j.issn:1671-3877.2000.02.014 |
万美亮, 邝炎华. 1998. 不同基因型甘蔗磷素吸收动力学特征研究初报. 华南农业大学学报, 19(2): 125-126. |
吴平, 印莉萍, 张立平, 等. 2001. 植物营养分子生理学. 北京: 科学出版社.
|
谢钰容, 周志春, 金国庆, 等. 2003. 马尾松不同种源磷素吸收动力学特征. 林业科学研究, 16(5): 58-553. |
谢钰容, 周志春, 金国庆, 等. 2004. 低磷胁迫下马尾松不同种源根系分泌的主要有机酸研究. 林业科学研究, 亚林所所庆专刊: 45-49. |
徐庆章. 1989. 小麦叶片酸性磷酸酯酶活性及缺磷对它的影响. 植物生理学通讯, 6: 20-23. |
严小龙, 黄志武, 卢仁骏, 等. 1992. 关于作物磷效率的遗传学研究. 土壤, 24(2): 102-105. |
周志春, 谢钰容, 金国庆, 等. 2003. 马尾松种源对磷肥的遗传反应及根际土壤营养差异. 林业科学, 39(6): 62-67. DOI:10.3321/j.issn:1001-7488.2003.06.010 |
Fox T R, Comerford N B. 1992. Rhizosphere phosphatase activity and phosphatase hydrolyzable organic phosphorus in two forested spodosols. Soil Biology and Biochemistry, 24(6): 579-583. DOI:10.1016/0038-0717(92)90083-A |
Goldstein A H, Baertlein D A, McDaniel R G. 1988. Phosphate starvation inducible metabolism in L. esculeutum. Ⅱ. The excreted phosphate starvation inducible APase of tomato. Plant Physiology, 87: 716-720. DOI:10.1104/pp.87.3.716 |
Hirata H, Hisaka H, Hirata A. 1982. Effect of phosphorus and potassium deficiency treatment on roots secration of wheat and rice seedlings. Journal of Plant Nutrition and Soil Science, 28: 543-552. DOI:10.1080/00380768.1982.10432394 |
Kummerva M. 1986. Localization of acid phosphatase in maize root under phosphorus deficiency. Biologia Plantarum, 28: 270-274. DOI:10.1007/BF02902291 |
Lefebvre D D, Duff S M G, File C, et al. 1990. Response to Phosphate deprivation in Brassica nigra suspension cells. Plant Physiology, 93(2): 504-511. DOI:10.1104/pp.93.2.504 |
Mclanchlan K D. 1980. Acid phosphatase activity of intact roots and phosphorus nutrition in plants. Ⅰ. Assay conditions and phosphatase activity. Australian Jorunal of Agricultural Research, 31: 429-440. DOI:10.1071/AR9800429 |