 2005, Vol. 41
2005, Vol. 41文章信息
- 黄荣凤, 张国盛, 鲍甫成, 王林和, 刘海东.
- Huang Rongfeng, Zhang Guosheng, Bao Fucheng, Wang Linhe, Liu Haidong.
- 毛乌素沙地臭柏年轮生长动态的研究
- Dynamic Analysis on Tree Ring Growth of Sabina vulgaris Grown in Mu Us Sandland
- 林业科学, 2005, 41(2): 117-122.
- Scientia Silvae Sinicae, 2005, 41(2): 117-122.
-
文章历史
- 收稿日期:2003-03-24
-
作者相关文章


2. 内蒙古农业大学 呼和浩特 010018
2. Inner Mongolia Agricultural University Huhhot 010018
树木的生长是由树木自身的遗传特性,以及立地条件、气候条件等环境因子所决定的。在适宜的环境条件下,温带的树木一般每年只形成一个生长轮,因此称之为年轮。但当生态环境条件恶劣,形成层活动受到抑制时,树木自身固有的遗传特性不能充分表现,因而有可能导致年轮畸形,出现伪年轮或不连续年轮。在相同立地条件下,树木的年轮生长则主要取决于气候因子的变化,特别是在干旱地区,树木的年轮生长主要受气候因子的制约。气候条件好,尤其是降雨量多的年份,树木的形成层活动旺盛,年轮就宽;相反气候条件不好的年份,形成层活动受到抑制,不仅年轮变窄,而且容易形成伪年轮(Fritts, 1976; Cook et al., 1977)。因此,研究树木年轮动态是把握树木生长与环境的关系,了解植物群落发生、发展和演替过程的重要手段。
臭柏(Sabina vulgaris)是沙漠地区重要的树种,对干旱、半干旱气候条件以及沙土基质环境有着很强的适应能力。在毛乌素沙地,臭柏是唯一的常绿针叶建群树种,主要分布在固定、半固定沙丘及丘间低地上,形成单优势的群落或明显的纯林(张国盛等,2001;Hirobe et al., 2001)。目前关于臭柏的生理生态学特性、更新环境、人工造林等的研究报道很多(张国盛等,1997;1999a;1999b;王林和等,1998a;1998b;2002;山中典和等,2001;温国胜等,2002),但还未见到有关天然臭柏年轮生长动态及其与环境关系的研究报道。本研究通过对毛乌素沙地天然臭柏年轮形态特征、年轮生长动态的分析,探讨了臭柏群落的起源以及臭柏年轮形成与环境的关系,目的是为毛乌素沙地臭柏群落的发生、发展、演替等生态学研究提供科学依据。
1 采样地概况和研究方法 1.1 样木与采样地概况以毛乌素沙地天然臭柏为试验材料。采样地点位于内蒙古鄂尔多斯市乌审旗图克苏木境内毛乌素沙地研究开发中心北试验区和图克臭柏保护区的天然臭柏分布区。地理位置为38°57′—39°01′N,109°02′—109°17′E,处于毛乌素沙地的中心。该地区属温带大陆性半干旱气候,年均气温6.5 ℃,年降水量为140~500 mm,降水量的年度变化大,且降水主要集中在7—9月,约占年降雨量的60%~70%;年均蒸发量2 300 mm。风大沙多,土壤为风沙土。地下水较丰富,滩地浅层地下水埋深小于3 m。地貌类型有滩地约25%;流动沙丘约占65%;固定、半固定沙丘约占10%,沙丘较为平缓,相对高度小于10 m。
北试验区的臭柏群落与油蒿(Artemisia ordosica)群落镶嵌分布,以成年期臭柏为主,主要分布于滩地边缘及沙丘顶部,生长状况良好,覆盖度为75%~90%;该臭柏分布区已无天然更新环境,几乎没有天然更新苗。臭柏保护区的臭柏群落从幼年期的更新苗到成年期的皆有,成年期臭柏主要分布于固定沙丘上,生长良好,覆盖度80%~95%;臭柏的天然更新只发生在有季节性积水的滩地边缘的乌柳(Salix cheilophila)、沙棘(Hippophae rhamnoides)、沙柳(Salix psammophila)灌丛下,幼年期的臭柏与乌柳、沙棘、沙柳混生,生长状况良好。
1.2 研究方法从北试验区的滩地臭柏灌丛中选择3株、沙丘顶部的臭柏灌丛中选择6株,从臭柏保护区的滩地臭柏灌丛中选择2株,共11株作为研究对象。每株选择1条生长良好的匍匐茎,在匍匐茎基部(地表面处)及距基部20 cm处各截取一个圆盘,作为年轮分析材料。其中,对北试验区沙丘顶部样木1条、臭柏保护区样木1条,由基部开始每隔20 cm截取一个圆盘,作为树干解析用试样。
对截取的圆盘进行磨光处理后摄影。在实体解剖镜下观察年轮特征,根据生长轮是否呈连续的圆环及不同断面生长轮的宽度变化判断伪年轮,并统计伪年轮出现的频率。
取通过髓心的木条,软化处理后分成8 mm×16 mm的平行四边形连续木块,所有的木块在HM-400滑走切片机上切成20 μm厚的横切面切片,用番红染色,系列酒精脱水,树脂胶封固,制成连续永久切片。在光学显微镜下进行显微摄影,结合伪年轮的判断结果,在放大20倍的连续照片上标记伪年轮并测量年轮宽度。照片测量值和实测值间的误差在2%以下。
2 结果与讨论 2.1 年轮形态特征和年龄确定臭柏群落包含水平生长的匍匐枝和垂直生长的直立枝。沙丘顶部臭柏受飞沙的影响大,经常发生枝条覆沙现象,容易形成不定根,主要以水平生长的匍匐枝为主;滩地臭柏由于水分充足,群落中直立枝相对比较多(王林和等,1998b)。对沙丘顶部和滩地臭柏观察发现沙丘顶部臭柏匍匐茎的偏心生长明显(图版Ⅰ-1),基部圆盘的平均偏心率(长径与短径比)为140.36%(表 1),常形成众多不连续生长轮或伪年轮,伪年轮与正常生长轮相交、闭合或呈不连续状(图版Ⅰ-3,4),且年轮窄,心材与边材区分较明显,基部心材比例较大;滩地臭柏匍匐茎的偏心生长不明显(图版Ⅰ-2),基部圆盘的平均偏心率为105.22%(表 1),没有发现不连续年轮,年轮宽度也较沙丘顶部臭柏宽,心材比例较小。大多数臭柏匍匐茎下部,即接近地面一侧的年轮较上部宽。

|
图版Ⅰ Plate Ⅰ |
|
|

树干或枝条倾斜生长时,由于重力作用的刺激引起植物激素分布的变化,使树干或枝条上部和下部的形成层活性产生差异,引起树木偏心生,形成应力木。针叶树的应力木在倾斜部位的下侧,即受压侧形成,因而称之为应压木(古野毅等,1994)。从臭柏的偏心形成部位看,与一般针叶树应压木的形成部位是一致的,但臭柏的偏心生长是否是由于重力作用引起,年轮较宽一侧是否是应压木,还需通过细胞形态观察及化学分析等手段进一步研究。
臭柏匍匐茎的生长轮清晰,早材占全部年轮的绝大部分,晚材带甚窄,由数列厚壁的长方形或圆形管胞构成,早材至晚材渐变(图版Ⅰ-5,6)。通常针叶树种晚材的管胞壁明显加厚(古野毅等,1994),而毛乌素沙地臭柏的管胞壁加厚现象在年轮形成初期、中期和后期都有可能出现,位置不确定(图版Ⅰ-4,5,6),这种现象是重力引起的应压木特征的表现,还是生长季内的气候因子多变,以及匍匐茎各个方向的微气候差异在树木年轮形成过程中的反映,尚需通过研究确定。
年轮特征分析结果如表 2所示,本研究所采集的滩地臭柏样木中40年生以上的臭柏有3株,年轮数最大的匍匐茎为50年生,伪年轮的发生频率为1.8%。沙丘顶部臭柏中40年生以上的有6株,年轮数最大的匍匐茎为123年生,伪年轮的发生频率高达12.07%。由此结果可以断定分布在固定沙丘顶部的天然臭柏群落至少有123年以上。
|
|
干旱沙漠温度变化大,降水不均衡。当生长季出现持续干旱或高温时,都会使树木暂时处于休眠状态,环境条件恢复后又重新开始生长,因此一年内可能形成2个或2个以上的生长轮,即伪年轮(刘宁,1994)。王炜等(2001)对干旱区梭梭(Haloxylon ammodendron)的年轮动态研究中也发现梭梭常有一年形成2个生长轮的现象,伪年轮的发生频率达37.5%。这可能是由于梭梭是古地中海起源的植物,具有夏季休眠的习性造成的。从沙丘顶部臭柏和滩地臭柏的年轮特征可以看出,臭柏的年轮形态、以及伪年轮的出现与土壤水分条件密切相关。
2.2 年轮生长动态 2.2.1 径向生长动态臭柏匍匐茎的树干解析结果(图 1)表明,滩地臭柏的偏心生长不明显,干形呈圆锥形;沙丘顶部臭柏的年轮表现出明显的偏心生长,贴近地面一侧的干形呈对数曲线形。由于臭柏匍匐茎上部的年轮生长缓慢,年轮宽度的变化不显著,所以本研究中径向动态分析统一使用下部的年轮测定数据。

|
图 1 臭柏的树干解析 Fig. 1 The stem parse of S. vulgaris A.沙丘顶部Top of dune;B.滩地Lowland. |
图 2是臭柏匍匐茎基部20 cm处的平均年轮宽度随年轮数的变化过程。生长于沙丘顶部的臭柏初期生长缓慢,年生长量在1 mm以下,此时期持续约14 a左右,之后径向生长有所增加,生长的高峰期出现在15~65 a之间,随后生长速度逐渐减慢。年平均生长量为0.94 mm,最大年生长量达2 mm(表 2)。整个生长过程中的年生长量变化很大,变异系数为72.45%,但看不出明显的峰值。

|
图 2 臭柏匍匐茎基部20 cm处年轮宽度的径向变异 Fig. 2 Variation of mean ring width at 20 cm from stolon base of S. vulgaris |
滩地天然更新的臭柏初期生长也慢,但年生长量的增长速度很快,5年生左右时年生长量达1 mm以上,35年时年生长量达到最大值4.5 mm,高峰期持续时期较沙丘顶部臭柏短,有明显的峰值,年平均生长量为1.51 mm,是沙丘顶部臭柏的1.6倍。生长量的年度间变化较沙丘顶部小,变异系数为52.96%。这一结果与毛乌素沙地小叶杨(Populus simonii)和北京杨(Populus × beijingensis)的径生长动态(Huang et al., 2000)相似。
树木的直径生长过程,可划分为幼年期、成年期和老年期3个阶段。一般幼年期持续时间较短,整个生长过程的累积生长曲线具有S型特征(Husch et al., 1972)。图 3是臭柏的径向累积生长曲线。沙丘顶部臭柏没有表现出一般树木所具有的阶段性生长的特性,整个生长过程S型特征不明显。滩地臭柏的S型特性也不明显。

|
图 3 臭柏的径向生长过程 Fig. 3 Radial growing process of S. vulgaris A.沙丘顶部Top of dune;B.滩地Lowland. |
干旱、半干旱地区的主要气候特点是温度和年降水量变化大,且不稳定。其中,水是制约这些地区植物生长的关键生态因子。毛乌素沙地处于半干旱气候带,沙丘顶部生长的臭柏表现出了幼年期生长缓慢且持续时间长、整个生长过程径向生长的变异系数大等特性,这与生长季水分不足、气候因子变化大有直接关系。可以认为,臭柏径向生长量的变化,是干旱地区的气候环境变化在树木上的反映。
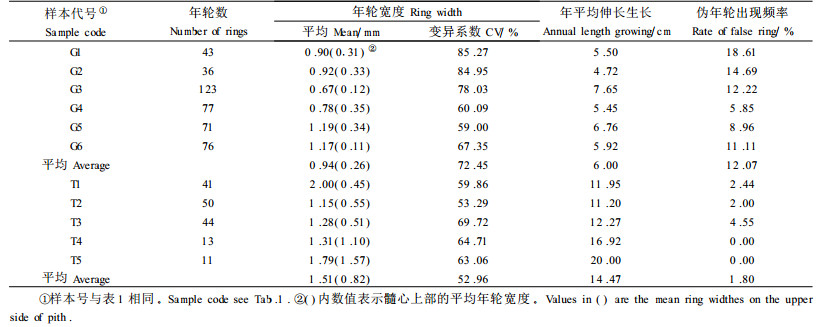
2.2.2 匍匐茎伸长生长动态从臭柏匍匐茎的伸长生长过程(图 4)可以看出,沙丘顶部臭柏的初期伸长生长缓慢,20龄时匍匐茎长仅为40 cm,37龄时匍匐茎的长度为120 cm,37龄之后生长迅速增加,37~43龄的6 a间伸长生长量达90 cm。沙丘顶部臭柏的整个伸长生长过程趋于指数曲线形增长,曲线方程为:

|
图 4 臭柏匍匐茎的伸长生长动态及拟合曲线 Fig. 4 Length growing dynamic and the fitting curve of stolon of S. vulgaris A.沙丘顶部Top of dune;B.滩地Lowland. |
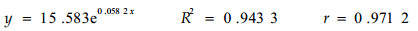
|
滩地臭柏匍匐茎伸长生长迅速,13龄时匍匐茎的长度达220 cm。滩地臭柏的整个伸长生长过程中,随着年龄的增加,伸长生长呈线形增长,线形方程为:
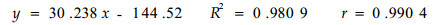
|
在上述2式中:x为匍匐茎的年龄,y为匍匐茎的长度,R2为决定系数,r为相关系数。
回归分析结果表明,臭柏的匍匐茎达到相同的长度时,沙丘顶部臭柏匍匐茎的长度与年龄之间存在着显著的非线性相关关系,而滩地臭柏的匍匐茎长度与年龄之间存在着显著的线性相关关系。2种立地条件下,臭柏伸长生长过程的差异说明水分条件与匍匐茎的伸长生长密切相关,水分不足时,臭柏的伸长生长受到抑制,特别是在匍匐茎伸长生长初期,这种抑制作用表现尤为明显。
臭柏匍匐茎的伸长生长使臭柏灌丛不断扩展,在毛乌素沙地形成郁闭度很大的臭柏群落。虽很多臭柏灌丛边缘生长旺盛并不断扩展,而中心部位生长缓慢甚至死亡。对臭柏灌丛不同部位叶面光合测定的结果表明,灌丛中心部位叶面的光合速率仅为边缘的1/5~1/6,灌丛的中心出现了生理衰退现象(温国胜等, 2002)。本研究中年轮分析的结果,沙丘顶部臭柏的匍匐茎进入衰退期时的平均年龄为65 a。
3 结论沙丘顶部和滩地臭柏的年轮形态明显不同。沙丘顶部臭柏偏心生长明显,长径与短径比为140.36%,伪年轮的发生频率为12.07%。滩地臭柏偏心生长不明显,长径与短径比为105.22%,伪年轮的发生频率为1.80%。根据匍匐茎的年轮数推断,沙丘顶部臭柏群落的年龄至少有123 a以上;滩地臭柏的年龄有50 a以上。
沙丘顶部臭柏径向生长缓慢,生长过程中没有明显的峰值,在65 a左右进入衰老期,年平均生长量为0.94 mm,累积生长曲线的S型特征不明显。滩地臭柏的径生长高峰出现在35 a,平均生长量为1.51 mm,是沙丘顶部臭柏的1.6倍,累积生长曲线的S型特征也不明显。沙丘顶部臭柏的匍匐茎长度生长与年龄之间表现出指数函数关系,而滩地臭柏的匍匐茎长度与年龄之间存在着显著的线性相关关系。
在地势较高的沙丘顶部生长的臭柏不仅容易形成伪年轮,而且径向生长和伸长生长均比滩地臭柏慢。这2种立地条件下,臭柏的年轮形态和伪年轮出现频率的不同,以及臭柏的径向生长和伸长生长过程的差异,说明水分条件与匍匐茎的年轮形成和生长速度密切相关。在干旱环境下,地下水是树木生长所需水分的主要来源,所以在毛乌素沙地进行臭柏的更新造林时,要充分利用丘间低地和滩地良好的水分条件,以保证更新造林的成功。
古野毅, 泽边攻. 1994. 组织と材质. 大津: 海青社, 21-32.
|
刘宁. 1994. 年轮及伪年轮的形成与环境的关系. 生物学通报, 29(9): 15. |
山中典和, 张国盛, 王林和, 等. 2001.  .日本绿化工学会志, 26(1): 120-124 .日本绿化工学会志, 26(1): 120-124
|
王林和, 张国盛, 董智. 1998a. 毛乌素沙地臭柏种子产量及更新的初步研究. 林业科学, 34(6): 105-112. |
王林和, 董智, 张国盛. 1998b. 毛乌素沙地天然臭柏群落新梢生长规律的研究. 内蒙古林学院学报, 20(3): 15-21. |
王林和, 刘美珍, 张国盛, 等. 2002. 毛乌素沙地臭柏不定根发生特性的研究. 林业科学, 38(5): 156-159. DOI:10.3321/j.issn:1001-7488.2002.05.026 |
王炜, 梁存柱, 朱宗元, 等. 2001. 梭梭年轮测定方法及生长动态的研究. 干旱区资源与环境, 15(1): 67-74. |
温国胜, 王林和, 吉川贤. 2002.  .日本绿化工学会志, 27(3): 526-532 .日本绿化工学会志, 27(3): 526-532
|
张国盛, 董智, 王林和. 1997. 臭柏生物生态学特性及生长繁殖的研究. 内蒙古林学院学报, 19(2): 69-75. |
张国盛, 王林和, 李玉灵, 等. 1999a. 毛乌素沙地臭柏根系分布及根量. 中国沙漠, 19(4): 378-383. |
张国盛, 李玉灵, 王林和, 等. 1999b. 半干旱地区臭柏造林初步研究. 内蒙古林学院学报, 21(1): 21-25. |
张国盛, 高润宏, 王林和, 等. 2001. 毛乌素沙地臭柏群落结构和生物多样性组成研究. 内蒙古农业大学学报, 22(4): 88-91. |
Cook E R, Jacoby G C. 1997. Tree-ring-drought relationships in the Hudson Valley, New York. Science, 198: 399-401. |
Fritts H C. 1976. Tree rings and climate. New York: Acadrmic Press, 377-383.
|
Hirobe M, Ohte N, Karasawa N, et al. 2001. Plant species effect on the spatial patterns of soil properties in the Mu-us desert ecosystem, Inner Mongolia, China. Plant and Soil, 234: 195-205. DOI:10.1023/A:1017943030924 |
Huang R, Furukawa I. 2000. Correlation between climatic factors and radial growth of Populus spp. planted in a semiarid region in China. Mokuzai gakkaishi, 46: 363-367. |
Husch B, Miller C I, Beers T W. 1972. Forest mensuration. New York: Ronald.
|