 2005, Vol. 41
2005, Vol. 41文章信息
- 罗建勋, 顾万春.
- Luo Jianxun, Gu Wanchun.
- 云杉天然群体表型多样性研究
- Study on Phenotypic Diversity of Natural Population in Picea asperata
- 林业科学, 2005, 41(2): 66-73.
- Scientia Silvae Sinicae, 2005, 41(2): 66-73.
-
文章历史
- 收稿日期:2004-03-02
-
作者相关文章



中国是世界云杉属种类分布最多的国家,有16种及变种。四川有13个种及变种,且自然分布广泛。云杉(Picea asperata)是川西北部、甘肃东南部、陕西西南部和青海东部亚高山地区森林演替的顶级群落和重要的工业用材树种,也是国内生长最快的云杉属树种(罗建勋等,2001)。
由于历史原因,云杉天然林大多数被采伐,有的遗传资源已经丢失。要对云杉天然群体进行科学的保护和合理利用,必须弄清种内群体的遗传多样性。表型多样性是遗传多样性研究的重要内容,利用表型性状研究群体的遗传多样性具有简便、快速和节省费用等优点,至今仍然是重要而且有效的(顾万春等,1998;罗建勋等,2004;李文英等,2002)。有关云杉属植物表型多样性研究较少,且这些研究都限于局部分布区的有限群体,取样和研究性状也有限(罗建勋等,2003;江洪,1992;李长喜,1988:Yeh et al., 1980;Khalil, 1984; Kurakin, 1990)。本文报道云杉表型多样性研究结果,主要目的是了解不同地理区域云杉的表型变异程度,揭示其表型变异规律。
1 材料与方法 1.1 群体选择与试验材料采集在云杉自然分布区内选择有代表性10个群体,基本网罗了该树种的整个自然分布区(图 1),且能反映出分布区的特点。各群体的地理生态因子见表 1。群体内单株选择条件参见罗建勋等(2003),每群体选取30个单株进行野外调查和采样,株间距离50 m以上。在每个单株树冠中上部南向2年生枝采集40~60个成熟球果,同时采集2年生针叶40枚。

|
图 1 云杉自然分布图 Fig. 1 The natural distribution of P.asperata |
借鉴Falkenhagen(1978),Putenikhin(1997)和Li等(1997)云杉属植物表型测定方法测量所有表型性状。用直尺测定球果长度,针叶长度、宽度(中央),种鳞长度、宽度(最宽端),种翅长度、宽度(最宽端)和种子长、宽度(最宽端),用游标卡尺测定球果径(中央直径)。每群体测定30单株(家系),每单株随机测定30个球果(测量精度为0.01 cm),30枚针叶(测量精度为0.01 cm)。从上述30个球果的中部随机各取一个完整种鳞(种翅)完成性状测定(测量精度为0.01 cm),球果干燥脱粒后测定上述球果干质量(测量精度为0.01 g),群体分单株脱粒净种后,每单株随机取30粒种子,测定每粒种子的长度和宽度(测量精度为0.01 cm),每单株随机取100粒种子称重,并换算成千粒重(测量精度0.01 g),每家系重复30次。
|
|

对各性状采用巢式设计方差分析,群体间表型分化系数(VST)的定义和分析见葛颂等(1988)。EGA的求算参见顾万春等(1997)。
2 结果与分析 2.1 云杉群体间的形态变异特征云杉群体间和群体内表型性状的变异均方及F值见表 2。经F检验,球果、针叶、种鳞、种翅和种子的17个表型性状在群体间和群体内都存在显著差异。17个表型性状均值及均值的多重比较见表 3。从表 3可以看出,云杉球果的形态特征在群体间存在显著差异,球果长度、球果径、球果长度/径和球果干质量的变异特征都是南部群体小金(XJ)最大,北部卓尼群体(ZN)最小。针叶长度是西北部阿坝群体(AB)最短,东北部大录群体(DL)最长。针叶宽度最小为铁布(TB)群体,最大为北部群体卓尼(ZN),针叶长度/宽度最大值为南部群体小金(XJ),最小值为北部卓尼群体(ZN)。种鳞长度和宽度的变异特征相近,均值最大为南部小金群体(XJ),最小为北部卓尼群体(ZN);种鳞长度/宽度均值最大群体仍为南部小金群体(XJ),而最小值却是巴西群体(BX)。群体间种翅长度均值最长为南端小金(XJ)群体,最小为北部卓尼群体(ZN);种翅宽度均值最大群体为川盘(CP),最小群体为卓尼(ZN)、阿坝(AB)和巴西(BX)群体,种翅长度/宽度均值最大群体为林坡(LP),均值最小为北部卓尼群体(ZN)。群体间种子长度均值最大为南端小金群体(XJ),最小为北部卓尼群体(ZN)和西北部阿坝群体(AB)。种子宽度均值最大群体为大录群体(DL),最小为阿坝(AB)和巴西(BX)群体。综上所述,云杉绝大多数表型性状呈现以纬度为主的单向变异趋势,自然分布区从北到南,随着纬度的降低,其群体最高分布海拔呈不断上升的趋势(刘庆,2002),表型性状呈现由小变大的趋势。
|
|
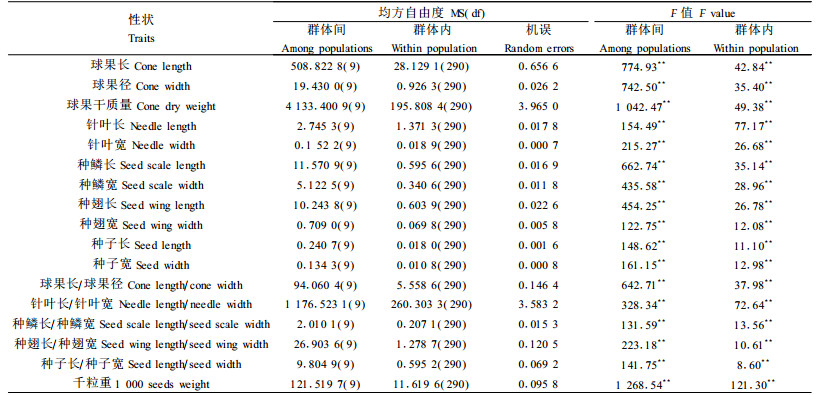
|
|

用变异系数表示性状值离散性特征,变异系数越大,则形状值离散程度越大。云杉10个群体表型性状的变异系数见表 4。从表 4可以看出,云杉球果、针叶、种鳞、种翅和种子5个生物学性状的平均变异系数(CV)分别为19.14%、26.46%、13.58%、19.78%和17.40%,说明种鳞性状较其他性状的稳定性高。在17个表型性状中球果干质量的变异系数最大(CV=34.10%)说明干质量较其他性状变异大;再有,球果、针叶、种翅和种子4个性状的形状指数比单个性状的变异系数大,球果长/径、针叶长/宽、种翅长/宽和种子长/宽的变异系数(CV)分别为14.88%、31.19%、21.70%和18.96%。仅有种鳞的形状指数的变异系数为最小(CV=11.99%),说明该性状的形状指数较单个性状稳定。从表 4还可看出,不同群体内每一性状的变异系数均有一定差异,说明不同群体的环境异质性导致了群体表型变异的差异。
|
|
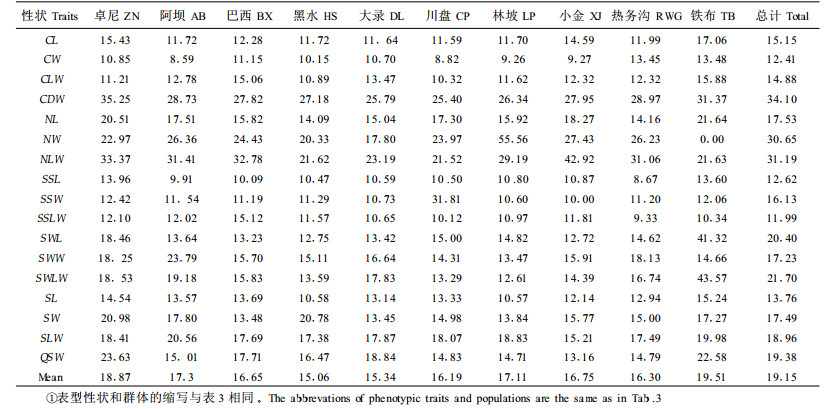
进一步对各群体变异系数的比较表明(表 4),北部卓尼(ZN)和铁布(TB)2群体的球果、种鳞、种翅和种子等表型性状的变异系数较大,平均变异系数为19.19%,说明这2个群体的表型多样性较丰富,而黑水(HS)和大录(DL)群体的变异系数较小,平均变异系数为15.20%,这2个群体的表型多样性较低。这可能与林分的起源有关,卓尼和铁布2群体均为天然纯林,而黑水和大录都为天然次生林。这也可能预示云杉北部边缘群体有较高的遗传变异。
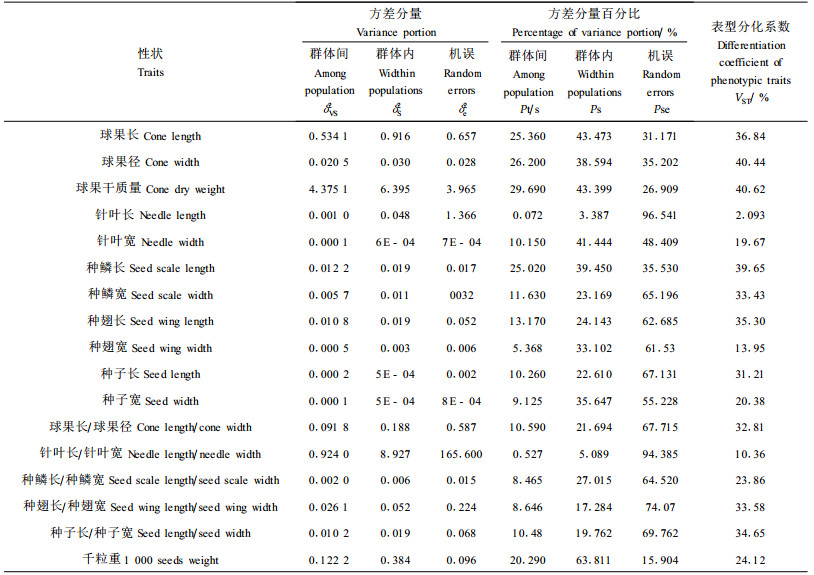
2.3 云杉天然群体间表型分化按巢式设计方差分量比组成了各方差分量占总变异的比例(表 5)、用群体间方差分量占总遗传变异(群体间群体内方差分量之和)的百分比表示群体间的表型分化系数VST。从表 5可以看出,球果、针叶、种鳞、种翅和种子性状VST的变异幅度为2.09%~40.62%,其中球果、种鳞、种翅和种子4性状的VST的均值分别为37.68%、32.31%、27.61%和28.75%,针叶性状的VST最小(10.70%)。球果、针叶、种鳞、种翅和种子17个表型性状的平均表型分化系数VST为30.99%,群体间变异(30.99%)小于群体内变异(69.01%),说明云杉天然群体内变异显著高于群体间变异。
|
|
利用欧式平均距离,采用类间平均连锁法对10个群体的球果、种鳞、种翅和种子表型数据进行聚类分析,结果见图 2。以欧氏距离5~10为阈值,10个群体明显分为5组,北部卓尼群体(ZN)和巴西群体(BX)聚为一组,南端的小金群体(XJ),北部铁布群体(TB)和西部阿坝群体(AB)均单独聚为一组,中心群体大录(DL)、川盘群体(CP)、黑水群体(HS),林坡群体(LP)和热务沟群体(RWG)明显聚为一组。进一步表明云杉边缘群体和中心群体的表型性状有明显差异。

|
图 2 云杉表型性状聚类树型图 Fig. 2 UPGMA cluster based on the phenotypic traits of ten populations in P.asperata |
对云杉球果、针叶、种鳞、种翅和种子17个性状平均值进行了相关分析与检验。结果表明,云杉球果长度与球果干质量、种鳞长度、种鳞宽度、种翅长度和种子千粒重5个性状呈极显著正相关,相关系数分别为0.831**、0.823**、0.904**、0.903**和0.841**:球果长/径与种翅长度呈极显著正相关(r=0.650**);球果干质量与种鳞长度、宽度和种翅长度呈极显著正相关,相关系数分别为0.919**、0.834**和0.810**;针叶长/宽与种翅长度呈极显著正相关(r=0.835**);种鳞长度和宽度呈极显著正相关(r=0.789**);种鳞长度与种翅长度也呈极显著正相关(r=0.865**);种鳞宽度与种翅长度、宽度和种子千粒重呈极显著正相关,相关系数分别为0.829**、0.772**和0.820**;种翅长度与种翅长/宽和种子长度呈极显著正相关,相关系数分别为0.712**和0.663**。综上所述,为提高云杉表型性状测定效率,今后测定种实性状时,球果长度、球果的径、种鳞的长度、种翅长度和种子千粒重应视为重要和关键表型性状。
2.6 云杉表型性状和采集点地理生态因子的相关关系对云杉球果、针叶、种鳞、种翅和种子的17个性状与采样地的地理生态因子进行了相关分析与检验。结果表明,云杉球果径与纬度呈极显著的负相关(r=-0.812**),种子长/宽与经度呈极显著负相关(r=-0.846**),从西向东,种子长度减小,宽度增加。球果干质量、种鳞长度和种鳞长/宽与生态梯度值呈极显著负相关,相关系数分别为-0.209**、-0.247**和-0.366**,针叶宽度与生态梯度值呈极显著正相关(r=0.262**)。球果干质量与纬度呈显著负相关(r=-0.681*),种翅长/种翅宽与经度呈显著负相关(r=-0.674*),种子宽度与经度呈显著正相关(r=0.641*),球果长度、种鳞长度、种鳞宽度和种翅长度与年均温呈显著正相关,相关系数分别为0.697*、0.715*、0.674*和0.759*。种子千粒重与1月均温和生态梯度值呈显著正相关,相关系数为0.663*和0.138*。种鳞宽度与7月均温呈显著正相关(r=0.636*),球果长/球果宽与针叶长/针叶宽及生态梯度值呈显著负相关,相关系数为-0.138*和-0.123*。
3 结论与讨论 3.1 云杉的表型变异极其丰富云杉种内存在着丰富的群体间、群体内变异。目前所见报道,云南松(Pinus yunnanensis)群体间的表型遗传变异较大(VST=36%),这主要是云南松为了适应云贵高原复杂多变的生态地理环境所致(陈少瑜;2001)。与其他云杉树种相比,云杉群体间的表型分化系数为中等偏高水平,高于西加云杉(Picea sitchensis)(VST=10.1%)(Yeh et al., 1980)和红皮云杉(Picea koraiensis)(VST=14.51%),低于黑云杉(Picea mariana)(VST=49.41%~57.04%)和白云杉(Picea glauca)(VST=50%)(张含国等,2003)。与其他针叶树种相比,云杉表型分化系数同长白落叶松(Larix olgensis)和白皮松(Pinus bungeana)接近,VST分别为26.2%和22.86%,高于花旗松(Pseudotsuga menziesii)、马尾松(Pinus massoniana)、北美短叶松(Pinus banksiana)和火炬松(Pinus taeda),VST分别为11.1%、6.44%、1.6%~18.9%和15%(张含国等,2003)。
3.2 云杉的表型变异呈梯度规律性Stebbbins(1950)认为,比起生长性状,黑云杉天然群体的花结构性状(球果和种子)受强的遗传控制。云杉球果性状以纬度梯度变异为主,随着纬度的增加,从南到北,球果的宽度逐渐变小,干质量逐渐变小。这与欧洲云杉的研究结果相异,Khalil(1984)认为欧洲云杉球果性状的变异以经度变异为主,与黑云杉研究结果也不同,Khalil(1985)认为球果性状变异的地理趋势不明显,球果长度、宽度和烘干质量及种子千粒重受强的遗传控制;云杉千粒重与一月均温呈显著正相关(r=0.663*),这与黑云杉的研究结果相反,Khalil(1984)认为黑云杉天然群体的种子千粒重受微弱的环境控制;云杉种鳞宽度与七月均温呈显著正相关(r=0.636*),这与Roche(1969)的研究结果亦相反,他们认为白云杉、黑云杉、西加云杉和恩氏云杉的种鳞的表型性状受强的遗传控制。也就是说,云杉球果、种鳞、种翅和种子性状的地理梯度变异都与温度有关,这与欧洲云杉的研究结果相同,研究结果认为温度是欧洲云杉地理变异的主要选择因子(Bergmann,1978)。
3.3 云杉遗传改良前景据云杉丰富的自然变异,可以推断云杉的改良前景是广阔的。云杉天然群体30.99%的变异存在于群体间,69.01%的变异存在于群体内,群体内的分化大大高于群体间。因此,在遗传改良工作中,在进行优良种源、优良群体选择和利用同时,应加大优良个体选择和利用的力度。因为我们需要的不是亲本的表现而是后代在新环境中的生长情况(Lines,1967)。
陈少瑜, 吴丽圆, 李江文, 等. 2001. 云南红豆杉天然种群遗传多样性研究. 林业科学, 37(5): 41-48. DOI:10.3321/j.issn:1001-7488.2001.05.008 |
顾万春, 李斌, 游应天. 1997. 生态梯度轴(EGA)区划林木育种区的研究. 生态学报, 17(2): 5-8. |
顾万春, 王棋, 游应天, 等. 1998. 森林遗传资源学概论. 北京: 中国科学技术出版社, 1-296.
|
葛颂, 王明庥, 陈岳武. 1988. 用同工酶研究马尾松群体的遗传结构. 林业科学, 24(4): 399-409. |
江洪. 1992. 云杉种群生态学. 北京: 中国林业出版社, 120-143.
|
罗建勋, 左林. 2001. 云杉人工林材性变异的初步研究. 西北农林科技大学学报, 29(3): 29-34. |
罗建勋, 李晓清, 孙鹏, 等. 2003. 云杉天然群体表型变异研究. 东北林业大学学报, 31(1): 9-11. DOI:10.3969/j.issn.1000-5382.2003.01.003 |
罗建勋, 顾万春. 2004. 云杉表型与同工酶遗传多样性研究进展. 林业科学研究, 17(2): 255-262. DOI:10.3321/j.issn:1001-1498.2004.02.020 |
李斌, 顾万春, 卢宝明. 2002. 白皮松天然群体种实性状表型多样性研究. 生物多样性, 10(2): 181-188. DOI:10.3321/j.issn:1005-0094.2002.02.008 |
李长喜. 1988. 林木天然群体表型变异研究概述. 林业科学研究, 1(6): 657-664. |
李文英, 顾万春. 2002. 栎类植物遗传多样性研究. 世界林业研究, 15(2): 42-49. DOI:10.3969/j.issn.1001-4241.2002.02.007 |
刘庆. 2002. 亚高山针叶林生态学研究. 成都: 四川大学出版社, 103-110.
|
张含国, 孙立夫, 韩继风, 等. 2003. 红皮云杉群体遗传多样性研究. 植物研究, 25(5): 224-229. |
Bergmann F. 1978. The allelic distribution at an acid phosphatase locus in Norway Spruce(Picea abies) along similar climatic gradients. Theoretical and Applied Genetics, 52(2): 57-64. DOI:10.1007/BF00281317 |
Falkenhagen E R. 1978. Multivariate classification in provenance research. Silvae Genetica, 27(1): 14-23. |
Khalil M A K. 1984. Genetic of cone morphology of black spruce (Picea mariana(Mill). B.S.P) in Newfoundland, Canada. Silvae Genetica, 33: 101-109. |
Khalil M A K. 1985. Genetic variation in estern white spruce (Picea glauca(Moench) Voss) populations. Can J For Res, 15: 444-452. DOI:10.1139/x85-071 |
Khalil M A K. 1984. Genetic variation in black spruce (Picea mariana(Mill) B.S.P.) in Newfoundland. Silvae Genetica, 24(4): 88-96. |
Kurakin B N. 1990. Variation in the number of cotyledons in seedling of spruce of different geographical origin. Lesnoe Khozyaistvo, (9): 39-40. |
Lines R. 1967. Standardization of methods for provenance research and testing. XIV.IUFRO-Kongress, section 22-AG22/24.672-718
|
Li Peng, Beaulieu J, Bousquet J. 1997. Genetic structure and patterns of genetic variation among populations in eastern White Spruce(Picea glauca). Canadian Journal of Forest Research, 27(2): 189-198. DOI:10.1139/x96-159 |
Putenikhin V P. 1997. Phenotype analysis of Picea obovata in the southern Urals population structure. Lesovedenie, (6): 37-49. |
Roche L. 1969. A genecological study of the genus Picea in British Columbia. New Phytol, 68: 505-554. DOI:10.1111/j.1469-8137.1969.tb06459.x |
Stebbins G L. 1950. Variation and evolution in plants. New York: Columbia Univ Press.
|
Yeh F C, EI-kassaby Y A. 1980. Enzyme variation in natural populations of Sitka spruce. Can J For Res, (10): 415-422. |