 2005, Vol. 41
2005, Vol. 41文章信息
- 郑淑霞, 上官周平.
- Zheng Shuxia, Shangguan Zhouping.
- 辽东栎叶片气孔密度及δ13C值的时空变异
- Tempo-Spatial Variations in the Stomatal Density andδ13C Value of Quercus liaotungensis Leaves
- 林业科学, 2005, 41(2): 30-36.
- Scientia Silvae Sinicae, 2005, 41(2): 30-36.
-
文章历史
- 收稿日期:2003-08-04
-
作者相关文章


随着全球人口剧增与工业化进程的加快,大气CO2浓度持续升高,导致全球气候环境发生明显的改变,这必然对陆地生态系统和植被分布产生很大的影响。植物叶面气孔密度对外界气候环境变化十分敏感(Uprety et al., 2002;贺新强等,1998),植物水分利用效率(WUE)是探明植物对于全球环境变化适应性以及预测全球变化影响的重要指标(郑凤英等,2001;Farquhar et al., 1989),植物体内稳定碳同位素特性已成为一种公认的分析不同生境下植物长期WUE变化的技术(孙谷畴等,1993;上官周平,2000;Warren et al., 2001;Condon et al., 2002)。因此用植物气孔密度和δ13C值来探讨植物对气候环境因子的适应性已成为目前植物生理生态学研究的热点之一(Uprety et al., 2002;Ziegler,1995;孙艳荣等,2002;Kurschner,2002)。
辽东栎(Quercus liaotungensis)是我国特有的树种,也是我国暖温带落叶阔叶林的主要优势树种之一,由于辽东栎既能适应偏冷潮湿气候,又具有耐温干气候的广生态幅特点,故在我国分布范围甚广,东北部到长白山北端,西北到清涧—安塞—志丹—吴旗一线,东界在朝鲜,西界到宁夏固原、青海循化(朱志诚,1982;金明仕,1987)。以辽东栎为优势种或建群种的森林群落稳定性高,其对区域植被的建成和群落演替有较大影响(王巍等,1999)。目前对辽东栎的种群结构与分布、生理生态、生长特性等方面的研究较多(林舜华等,1997;王巍等,1999;孙书存等,1999;高贤明等,2001;吴晓莆等,2002),但在大的时空尺度上研究辽东栎对气候环境变化的适应问题还较为缺乏。本文以不同植被区域的辽东栎为对象,利用数码图像显微镜处理系统和稳定性碳同位素测定技术,研究20世纪30—80年代辽东栎气孔密度及叶片δ13C值的时空变异规律,以揭示不同植被区域辽东栎对气候变化的生态适应对策,其结果将对森林植被特别是退化森林生态系统的恢复与重建具有重要意义。
1 材料与方法 1.1 材料的选取辽东栎植物样品取自中国科学院西北植物研究所标本馆,该标本馆保存有自20世纪20年代的约55万份植物标本。由于植物标本上详细记录了采集时间、地点以及植物生长的基本环境条件如海拔高度、阳坡阴坡及采集时的林冠高度等,这就为保证选取材料的一致性,尽量减小样本间的误差,提供了可靠的依据。本试验在选样时充分考虑了所选植物标本采样时间、地点的代表性与典型性。所选辽东栎标本大多数采自海拔1 000~1 500 m(青藏高原除外)的阴坡、半阴坡,林冠下部(距地面2~3 m),每份标本上选择生长状况良好的典型叶片(健康、成熟、叶脉分明且无虫叮咬痕迹)。取样原则为从20世纪30—80年代分30、50、80年代3个年代取样,取样年份、取样量及取样地区均视标本藏量而定,一般每一年代1~2份,每份样品由同一年代不同年份(1~2)不同地点(2~3个)的多份标本叶片组成,共计60份测定样品,240余份植物标本。所取植物样品采样时间一般为6—10月,为大多数植物的生长季。根据辽东栎在各地区的分布,研究地区西起101°18′E(四川阿坝藏族自治州),东至121°36′E(山东牟平县),南起25°12′N(贵州望漠县),北至42°36′N(吉林安图县),穿越吉林、河北、山西、河南、山东、江苏、陕西、宁夏、甘肃、四川、贵州、青海等12个省(自治区)。但由于受标本馆藏量所限及辽东栎这一物种不同地区分布面积的差异,有些地区样品可达十几份,如陕北黄龙山、甘肃天水等,而有些地区样品仅一两份,如四川阿坝藏族自治州、贵州望漠县。依据中国植被区域划分方法(吴征镒,1980),所选植物样品主要分布于温带针阔叶混交林、暖温带落叶阔叶林、亚热带常绿阔叶林、温带草原、青藏高原高寒植被等5个区域,其中多数样品来自暖温带落叶阔叶林区域。
1.2 植物叶片气孔密度的观测 1.2.1 临时装片的制备为防止气孔变形,采用印迹法制片。从每份标本上选取3片健康、成熟的叶片(较大,且无虫叮咬痕迹),用脱脂棉蘸酒精轻轻擦拭其下表皮灰尘,然后在下表皮中部靠近主脉的两侧快速涂上一层薄薄的透明指甲油,约1~1.5 cm2,待其风干结成膜后,用贴有两面粘性透明胶带的载玻片压在叶片上,然后轻轻剥下叶片,即把所有叶表皮膜的指甲油层粘在透明胶带上,制成表皮印迹后的载玻片用中性树胶封片,制成临时装片,于数码图像显微镜下进行观测。
1.2.2 气孔密度的观测利用具有130万像素的DMB5-223IPL数码显微镜摄像系统(Motic Digital Imaging,中国)观测。MoticTek模块能够将捕捉的图像高速导入电脑,图像分辨率为1 280×1 024活动像素,图像处理系统采用Motic Images Advanced(3.0)软件,可自动显示测量结果,并直接导出*.xls文档,进行数据分析。
每份标本各制3个临时装片于40倍数码显微镜下拍照观测。数码图像显微镜系统中的采集窗能够自动捕捉图像,MoticTek模块能够将捕捉的图像高速导入电脑并自动拍照,每一装片上随机选30个视野,即每个样品共拍60幅图片,计算每幅图片上的气孔个数,取平均值,除以图片面积,统计每mm2叶片上的气孔数,即气孔密度(stoma·mm-2),然后对数据进行统计分析。
1.3 植物叶片稳定性碳同位素组分(δ13C)值的测定δ13C值的测定在中国科学院南京土壤研究所土壤圈物质循环重点实验室进行,分析仪器为MAT-251质谱仪。从每份标本上选取1~2片健康、成熟叶片(较大,且无虫叮咬痕迹)磨碎,过80目筛后各取1 g,同一年代不同年份的多个地点的样品混合后制成供试样品。取处理好的样品3~5 mg封入真空的燃烧管,并加入催化剂和氧化剂,在850 ℃下气化,燃烧产生的CO2经结晶纯化后,测定δ13C值,以PDB(Pee Dee Belemnite)为标准,根据Farquhar等(1989)进行计算:
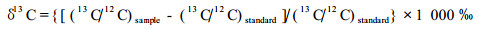
|
式中:(13C/12C)sample是植物叶片样品的13C/12C比率,(13C/12C)standard是测定过程中标准物质甘氨酸的13C/12C比率。计算的最终结果都以国际标准物质PDB为标准,分析误差≤0.2‰。
2 结果与分析 2.1 气孔密度和δ13C值的变化范围从20世纪30—80年代近50年中,随气候环境变化,全国不同区域中辽东栎叶片气孔密度变化范围为535~800 stoma·mm-2,平均值为703 stoma·mm-2,δ13C值变化范围为-28.47‰~-25.02‰,平均值为-26.83‰。就全国时空尺度来看,辽东栎叶片气孔密度和δ13C值的最大值均为暖温带落叶阔叶林区域中30年代植物样品,而最小值均为温带草原区域中80年代植物样品。
2.2 不同植被类型区域不同年代差异比较为了比较近50年中不同地区辽东栎叶片气孔密度和碳同位素组分分布的时空差异,选30年代、50年代及80年代中不同年份气孔密度和δ13C值的平均值进行比较(表 1,表 2)。将辽东栎样品较全的暖温带落叶阔叶林、亚热带常绿阔叶林、青藏高原高寒植被区域中30年代、50年代及80年代气孔密度和δ13C值作图比较(图 1),3个植被区域中,暖温带落叶阔叶林中辽东栎叶片气孔密度和δ13C值变化最大,其次为亚热带常绿阔叶林,青藏高原高寒植被区域变化最小。由20世纪30—80年代,暖温带落叶阔叶林区域中辽东栎叶片气孔密度和δ13C值持续降低较为明显;由20世纪30—50年代,亚热带常绿阔叶林区域中辽东栎叶片气孔密度呈增加趋势,而δ13C值呈降低趋势;青藏高原高寒植被区域中辽东栎叶片δ13C值呈增加趋势,但气孔密度几乎没有变化。
|
|

|
|


|
图 1 20世纪不同植被类型区域辽东栎叶片气孔密度(A)及δ13C值(B)变化 Fig. 1 The leaf stomatal density(A) and δ13C value(B) in leaf specimen of Q. liaotungensis collected from different vegetation region in the 20th century WTDB:暖温带落叶阔叶林区域Warm temperate deciduous broadleaf forest region;STEB:亚热带常绿阔叶林区域Subtropical evergreen-broadleaf forest region;TPAC:青藏高原高寒植被区域The Ti-betan Plateau alpine-cold vegetation region.下同。The same below. |
选辽东栎样品较全的暖温带落叶阔叶林区域中的甘肃、陕北、山西,比较从20世纪30—80年代近50年中,随气候环境变化,不同地区不同年代辽东栎叶片气孔密度和δ13C值的变化情况(图 2)。在近半个世纪中,不同地区辽东栎气孔密度对气候环境变化的反应有差异,山西辽东栎气孔密度呈明显下降趋势;而陕北和甘肃辽东栎气孔密度呈略微上升趋势,几乎没有变化。整体上,3个地区辽东栎不同年代气孔密度平均值的比较:山西(733 stoma·mm-2)>陕北(656 stoma·mm-2)>甘肃(599 stoma·mm-2)。在近半个世纪中,不同地区辽东栎叶片δ13C值均呈明显下降趋势,整体上,3个地区辽东栎不同年份δ13C值的平均值比较:山西(-25.61‰)>陕北(-26.80‰)>甘肃(-27.40‰)。

|
图 2 各个省份不同年份辽东栎叶片气孔密度(A)及δ13C值(B)变化 Fig. 2 The leaf stomatal density(A) and δ13C value(B)of Q. liaotungensis in different years each province |
将不同植被类型区域的辽东栎植物样品按30年代、50年代、80年代划分,比较3个年代不同植被类型区域中,辽东栎叶片气孔密度(图 3)和δ13C值的变化情况(图 4)。

|
图 3 20世纪30年代(A)、50年代(B)、80年代(C)不同植被类型区域辽东栎叶片气孔密度变化 Fig. 3 The leaf stomatal density of Q. liaotungensis in different vegetation region in 1930's(A)、1950's(B) and 1980's(C) TCBM:温带针阔叶混交林区域Temperate coniferous-broadleaf mixed forest region;TGR:温带草原区域Temperate grassland region.下同。The same below. |

|
图 4 20世纪30年代(A)、50年代(B)、80年代(C)不同植被类型区域辽东栎叶片δ13C值变化 Fig. 4 The leaf δ13C value of Q. liaotungensis in different vegetation region in 1930's(A)、1950's(B) and 1980's(C) |
随气候环境变化,20世纪30年代,暖温带落叶阔叶林、亚热带常绿阔叶林、青藏高原高寒植被3个区域中辽东栎叶片气孔密度依次递减,分别为800、713、686 stoma·mm-2。50年代,4个植被区域类型之间辽东栎叶片气孔密度差异不大,温带针阔叶混交林区域(776 stoma·mm-2)≈亚热带常绿阔叶林区域(775 stoma·mm-2)>暖温带落叶阔叶林区域(700 stoma·mm-2)≈青藏高原高寒植被区域(695 stoma·mm-2)。80年代,暖温带落叶阔叶林区域中的辽东栎气孔密度(643 stoma·mm-2)明显高于温带草原区域中的辽东栎气孔密度(535 stoma·mm-2)。
2.4.2 叶片δ13C值差异比较随气候环境变化,20世纪30年代,暖温带落叶阔叶林、亚热带常绿阔叶林、青藏高原高寒植被3个区域,辽东栎叶片δ13C值依次递减较明显,分别为-25.02‰、-26.05‰、-27.46‰。50年代,4个植被区域类型之间辽东栎叶片δ13C值差异不大,温带针阔叶混交林区域(-26.57‰)≈亚热带常绿阔叶林区域(-26.75‰)≈暖温带落叶阔叶林区域(-26.63‰)>青藏高原高寒植被区域(-27.10‰)。80年代,暖温带落叶阔叶林区域中的辽东栎叶片δ13C值(-27.41‰)明显高于温带草原区域中的辽东栎叶片δ13C值(-28.47‰)。
3 讨论目前,以植物标本为材料,研究植物气孔密度与气候环境变化的相关性的报道较多(Woodward,1987;Penuelas et al., 1990;贺新强等,1998;郑凤英等,2001)。Woodward(1987)利用1750年采集的8种温带树种的腊叶标本研究发现,与同种植物的气孔密度对比,由于大气CO2浓度升高60 μmol·mol-1,气孔密度下降了40%。贺新强等(1998)研究了中国特有的5种木本植物气孔密度在近一个世纪中随大气CO2浓度升高的变化情况,结果表明,杜仲(Eucommia ulmoides)、辽东栎、短柄枹栎(Q.glandulifera var. brevipetiolata)的气孔密度明显降低,青钱柳(Cyclocarya paliurus)的气孔密度降低不明显,而异叶榕(Ficus heteromorpha)的气孔密度变化无规律性。郑凤英等(2001)研究了亚热带气候区域内的狭叶山黄麻(Trema angustifolia)和光叶山黄麻(Trema cannabina)2种植物腊叶标本气孔密度在近一个世纪里的变化,发现从20世纪20年代到90年代,随大气CO2浓度升高,2种植物气孔密度分别降低28.1%和40.0%。上述研究结果只考虑了CO2浓度这一单因子变化,却没有考虑到其他环境因子的影响作用。
本试验中,从时间分布角度,20世纪30—80年代,辽东栎叶片气孔密度变化:暖温带落叶阔叶林区域中逐年递减;亚热带常绿阔叶林区域中呈增加趋势;而青藏高原高寒植被区域中几乎没有变化。这一结论与Woodward(1987)、贺新强等(1998)和郑凤英等(2001)的研究结果并不完全一致,这可能是因为研究物种、区域不同。森林植被是一个复杂的生态系统,不同植被区域中土壤水分、养分、气候环境因子(光照、温度、降水量、海拔高度等)差异很大,各个区域中起主导作用的关键影响因子也不同,导致了即使是同一种植物在不同的生存环境中,也会有不同的适应策略。植物气孔密度与物种本身特性有关,除受CO2浓度变化的影响外,还与其他环境因子(温度、降水、光照等)的变化有关。3个植被区域中亚热带常绿阔叶林区域年降水量充沛,年均温高,而青藏高原高寒植被区域相对于前2个区域,海拔较高,年降水稀少,年均温低,这种较为明显的气候差异可能是导致气孔密度变化趋势不同的主要原因。辽东栎气孔密度变化从空间分布角度看,20世纪30年代,暖温带落叶阔叶林、亚热带常绿阔叶林、青藏高原高寒植被3个区域辽东栎气孔密度依次递减;50年代,辽东栎气孔密度在4个植被区域之间产生波动,温带针阔叶混交林和亚热带常绿阔叶林2个区域中的辽东栎气孔密度高于暖温带落叶阔叶林和青藏高原高寒植被2个区域;80年代,辽东栎在由东到西分布的暖温带落叶阔叶林至温带草原区域,气孔密度呈降低趋势。暖温带落叶阔叶林区域中,山西、陕北、甘肃3个省份辽东栎气孔密度依次降低,也表明辽东栎在由东至西相对较小的空间分布上,气孔密度呈降低趋势,这可能与山西、陕北、甘肃3个省份的气候干旱程度依次加剧有关。
许多研究结果表明,植物叶片的δ13C值与其WUE呈一定程度的正相关(Farquhar et al., 1989;O'leary,1988)。由于δ13C值是由遗传控制的,在植物体内较为稳定,因而测定植物叶片δ13C值就可以判定植物WUE高低,但同时植物δ13C值的环境效应仍然存在,在多种气候因子的影响下,植物种内δ13C值差异可达3‰~5‰,进而影响植物的长期WUE(O'leary,1988;Ziegler, 1995)。Gunderson等(1993)对美国白栎(Quercus alba)实生苗在CO2提高下的光合、呼吸及WUE等生理指标的变化进行了研究,发现光合速率及WUE均明显提高。林舜华等(1997)研究发现暖温带落叶阔叶混交林中建群种辽东栎在CO2倍增下,WUE明显提高,为对照的179%。林植芳等(1995)测定了亚热带常绿季风阔叶林下2种建群树种荷木(Schima saperba)和黧蒴(Castanopsis fissa)在不同光照强度(100%、40%、16%)下的幼苗叶片的δ13C值,发现随光照强度降低,δ13C值降低,WUE也降低。上述研究结果也多从CO2浓度、光照等单因子变化方面考虑其对植物WUE的影响。严昌荣等(1998;2001)对北京山区暖温带落叶阔叶林中6种木本植物叶片δ13C值进行了测定和比较,结果表明不同植物叶片δ13C值受多种因子的影响,具有较大的种间差异及时空变化;在植物生长季(1995年5—9月),6种植物叶片的δ13C值从生长初期到生长末期逐渐降低;除荆条(Vitex negundo var. heterophylls)外,辽东栎、核桃楸(Juglans mandshurica)、山杏(Prunus armeniaca var. ansu)、大叶白蜡(Fraxinus rhynchophylla)和北京丁香(Syringa pekinensis)的δ13C值均呈下降趋势。辽东栎叶片的δ13C值随其灌层高度的增加而上升。
本试验中,从时间分布角度,由20世纪30—80年代,随气候环境变化,暖温带落叶阔叶林和亚热带常绿阔叶林区域中辽东栎叶片δ13C值均呈降低趋势;而青藏高原高寒植被区域中辽东栎叶片δ13C值呈增加趋势。说明在前2个区域中辽东栎WUE逐年下降,而青藏高原高寒植被区域中辽东栎WUE却在增加,这可能与青藏高原具有独特的气候地理条件(尤其是海拔较高,约为其他2个区域的3~4倍)有关。本研究结论中关于暖温带落叶阔叶林和亚热带常绿阔叶林区域中辽东栎叶片δ13C值变化与严昌荣等(2001)、林植芳等(1995)的观点基本一致,同时笔者也认为植物叶片δ13C值除了由物种自身的遗传因素控制外,其生存环境的差异也会对δ13C值产生较大的影响。本文也对辽东栎叶片δ13C值变化从空间分布角度进行了研究。20世纪30年代,暖温带落叶阔叶林、亚热带常绿阔叶林、青藏高原高寒植被3个区域中辽东栎叶片δ13C值依次递减,指示了其WUE也呈递减趋势。50年代,4个植被区域类型之间辽东栎叶片δ13C值差异不大,说明其对应的植物WUE比较接近。80年代,暖温带落叶阔叶林区域中的辽东栎叶片δ13C值明显高于温带草原区域,且暖温带落叶阔叶林区域中,山西、陕北、甘肃3个省份中辽东栎δ13C值依次降低明显,也表明辽东栎在由东至西相对较小的空间分布上WUE下降。造成δ13C值变化趋势不同的主要原因应该也是不同植被带气候条件上的明显差异。
由于不同植被区域中光照、温度、CO2浓度、水分、养分等气候环境因素差异很大,不同年代和不同区域中起主导作用的关键影响因子也不同,使得辽东栎在不同的生存环境中气孔密度和δ13C值变化不同,但从大的时空分布上仍呈现出一定的规律性。在CO2浓度持续升高这一相同的环境背景下,除个别区域外,辽东栎在从20世纪30—80年代,由暖温带落叶阔叶林至亚热带常绿阔叶林、青藏高原高寒植被的时空分布上,气孔密度和δ13C值均呈降低趋势,至于导致这一趋势的影响机制有待于进一步研究。辽东栎叶片气孔密度与δ13C值的时空变异规律的研究将对这一优势种群的营造与抚育具有重要意义。
高贤明, 王巍, 杜晓军, 等. 2001. 北京山区辽东栎林的径级结构、种群起源及生态学意义. 植物生态学报, 25(6): 673-678. DOI:10.3321/j.issn:1005-264X.2001.06.005 |
贺新强, 林月惠, 林金星, 等. 1998. 气孔密度与近一世纪CO2浓度变化的相关性研究. 科学通报, 43(8): 860-862. DOI:10.3321/j.issn:0023-074X.1998.08.017 |
金明仕J P著.文剑平译.1987.森林生态学.北京: 中国林业出版社, 324-326
|
林舜华, 项斌, 高雷明, 等. 1997. 辽东栎对大气CO2倍增的响应. 植物生态学报, 21(4): 297-303. DOI:10.3321/j.issn:1005-264X.1997.04.001 |
林植芳, 林桂珠, 孔国辉, 等. 1995. 生长光强对亚热带自然林两种木本植物稳定碳同位素比、细胞间CO2浓度和水分利用效率的影响. 热带亚热带植物学报, 3(2): 77-82. |
上官周平. 2000. 小麦13C分辨率和水分利用效率对氮素和水分的响应. 植物营养与肥料学报, 6(3): 345-348. DOI:10.3321/j.issn:1008-505X.2000.03.015 |
孙谷畴, 林植芳, 林桂珠. 1993. 亚热带人工松林13C/12C比率和水分利用效率. 应用生态学报, 4(3): 325-327. DOI:10.3321/j.issn:1001-9332.1993.03.003 |
孙书存, 陈灵芝. 1999. 不同生境中辽东栎的构型差异. 生态学报, 19(3): 358-364. |
孙艳荣, 穆治国, 崔海亭. 2002. 北京地区近70年来白皮松树轮纤维素的碳稳定同位素与气候变化. 海洋地质与第四纪地质, 22(4): 85-90. |
王巍, 刘灿然, 马克平, 等. 1999. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 41(4): 425-432. |
吴晓莆, 郑豫, 马克平. 2002. 北京东灵山地区辽东栎、大叶白蜡和五角枫种群分布格局与动态. 植物学报, 44(2): 212-223. DOI:10.3321/j.issn:1672-9072.2002.02.016 |
吴征镒. 1980. 中国植被. 北京: 科学出版社.
|
严昌荣, 韩兴国, 陈灵芝, 等. 1998. 暖温带落叶阔叶林主要植物叶片中δ13C值的种间差异及时空变化. 植物学报, 40(9): 853-859. DOI:10.3321/j.issn:1672-9072.1998.09.014 |
严昌荣, 韩兴国, 陈灵芝. 2001. 六种木本植物水分利用效率和其小生境关系研究. 生态学报, 21(11): 1952-1956. DOI:10.3321/j.issn:1000-0933.2001.11.028 |
郑凤英, 彭少麟, 赵平. 2001. 两种山黄麻属植物在近一世纪里气孔密度和潜在水分利用率的变化. 植物生态学报, 25(4): 405-409. DOI:10.3321/j.issn:1005-264X.2001.04.004 |
朱志诚. 1982. 秦岭和陕北黄土高原辽东栎林研究. 植物生态学与地植物学学报, 6(1): 95-104. |
Condon A G, Richards R A, Rebetzke G J, et al. 2002. Improving intrinsic water-use efficiency and crop yield. Crop Science, 42: 122-131. DOI:10.2135/cropsci2002.1220 |
Farquhar G D, Ehleringer J R, Hubick K T. 1989. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 40: 503-507. DOI:10.1146/annurev.pp.40.060189.002443 |
Gunderson C A, Norby R J, Wullschleger S D. 1993. Foliar gas exchange responses of two deciduous hardwoods during 3 year of growth in elevated CO2:no loss of photosynthetic enhancement. Plant, Cell and Environment, 16: 789-807. |
Kurschner W M. 2002. Carbon isotope composition of fossil leaves-revealing ecophysiological responses to past environmental change. New Phytologist, 155(2): 199-201. DOI:10.1046/j.1469-8137.2002.00461_2.x |
O'leary M H. 1988. Carbon isotopes in photosynthesis. BioScience, 38: 328-336. DOI:10.2307/1310735 |
Penuelas J, Matamala R. 1990. Changes in N and S leaf content, stomatal density and specific leaf area of 14 plant species during the last three centuries of CO2 increase. Journal of Experimental Botany, 41: 1119-1124. DOI:10.1093/jxb/41.9.1119 |
Uprety D C, Dwivedi N, Mohan J V. 2002. Effect of elevated carbon dioxide concentration on the stomatal parameters of rice cultivars. Photosynthetica, 40(2): 315-319. DOI:10.1023/A:1021322513770 |
Warren C R, McGrath J F, Adams M A. 2001. Water availability and carbon isotope discrimination in conifers. Oecologia, 127: 476-486. DOI:10.1007/s004420000609 |
Woodward F I. 1987. Stomatal numbers are sensitive to increases in CO2 from pre-industrial leaves. Nature, 327: 617-618. DOI:10.1038/327617a0 |
Ziegler H. 1995. Stable isotopes in plant physiology and ecology. In: Behnke H D, Lüttge U, Esser K, et al (eds). Progress in Botany. Vol. 56. Berlin, Heidelberg, Germany: Springer-Verlag, 1-24
|