 2005, Vol. 41
2005, Vol. 41文章信息
- 孙辉, 谢嘉穗, 唐亚.
- Sun Hui, Xie Jiasui, Tang Ya.
- 坡耕地等高固氮植物篱复合经营系统根系分布格局研究
- Distribution Patterns of Root System under Contour Hedgerow Intercropping on Slopeland in Dry Valley of the Jinsha River
- 林业科学, 2005, 41(2): 8-15.
- Scientia Silvae Sinicae, 2005, 41(2): 8-15.
-
文章历史
- 收稿日期:2003-04-02
-
作者相关文章



等高固氮植物篱农林复合经营模式即在坡耕地上沿等高线每隔一定距离种植双行固氮植物篱,植物篱中间的土地为作物种植带。在这个系统中植物篱用于防治水土流失,刈割枝叶为农作物提供绿肥或用作牲畜饲料,也可覆盖地表加强防治土壤侵蚀的效果。在热带地区对这种模式的土壤培肥效果(Kang et al., 1981; Lal, 1989)、土壤理化性质变化(Kang et al., 1990; Agus et al., 1997)、水土保持效果(Kiepe, 1995)、植物篱与农作物之间的相互关系等进行了较多研究(Mclntyre et al., 1997; Imo et al., 2000)。唐亚、孙辉等在亚热带四川金沙江流域干旱河谷利用等高固氮植物篱综合治理坡耕地等方面进行了较详细的研究(Tang et al., 2000;2003; 孙辉等, 1999;2001),并在固氮植物篱枝叶矿化过程、复合经营、提高土地经济效益方面取得了进展(孙辉等,2002a;2002b;2003;袁远亮等,2000)。但是,对于植物篱复合经营系统中各组分之间关系的争议较大,而农林复合经营各组分之间的竞争或协同关系很大程度上是由于地下部分的相互作用引起的(马秀玲等,1997;Lehmann et al., 1998)。我国过去10余年的研究、试验、示范和初步推广应用结果均表明,植物篱农林复合经营是一项适宜于我国亚热带山区特点的种植模式,具有广阔的应用前景(唐亚等,2001)。但目前对这一系统中根系研究方面的工作还不够。本文通过对亚热带干旱河谷坡地等高固氮植物篱-桑树(Morus alba)-脐橙(Citrus sinensis)这种复合经营系统的根系分布格局的研究,旨在探索南亚热带和干旱河谷的植物篱复合经营系统地下部分的相互关系。
1 研究区域概况试验在地处金沙江干暖河谷区的四川省宁南县等高固氮植物篱试验与示范园进行。示范园地处金沙江支流干暖河谷气候区,旱季雨季明显,年降雨量660~1 140 mm,平均915.1 mm,5—10月的雨季降雨量占全年的87.9%。实验区海拔1 400~1 485 m,坡向西南,坡度15~38°;土壤为褐红壤,建点时表土有机质含量11.1 g·kg-1,全氮含量0.53 g·kg-1。示范园于1994年建立,植物篱树种主要为新银合欢(Leucaena leucocephala)、山毛豆(Tephrosia candida)等。
本研究所涉及的新银合欢固氮植物篱建立于1994年,植物篱通过刈割将高度控制在0.4~1 m左右,刈割的新鲜枝叶作为绿肥覆盖在植物篱之间的作物带地表和脐橙根部,可以同时防治土壤侵蚀和减少地表蒸发,刈割枝叶量平均为15 000 kg·hm-2。作物带中脐橙于1996年种植,1999年投产。新银合欢植物篱为双行,植物篱宽度50 cm,相邻植物篱间距4 m。新银合欢植物篱中套种桑树,桑树间距为2 m;植物篱之间的作物带种植一行脐橙,脐橙株距为2 m。
2 研究方法根系研究方法采用收获法与根钻法相互结合。根钻直径20 cm,深10 cm,取样时水平中心位置分别为-1.7、-1.1、-0.5、0、0.5、1.1、1.7 m,土壤剖面深度中心为30、60和90 cm(取样位置如图 1所示)。5次重复。

|
图 1 坡地植物篱复合经营示意图 Fig. 1 Contour hedgerow intercropping system along slope |
采挖剖面(取样剖面)为植物篱下侧作物带中间至植物篱上侧作物带之间,顺坡长4.5 m×宽0.6 m×深1.0 m,层次为0~45、45~70、70~100 cm;沿坡面以植物篱为中心,-2.0~-1.4、-1.4~-0.8、-0.8~-0.2、-0.2~0.2、0.2~0.8、0.8~1.4、1.4~2.0 m分段采挖,如图 1所示(植物篱下侧为负)。3次重复。
根据新鲜根颜色的明显差异,将新银合欢、桑树和脐橙根系分别捡出,剔除死根。细根分级标准从<1 mm到<5 mm不等(张小全等,2001),一般认为<1 mm的细根为吸收根或吸水根(Brunner et al., 2002;马秀玲等,1997),本研究按直径≥1 mm和<1 mm将根分为2级。分别测量各层次各位置不同植物根数(≥1 mm根的根数为该土层的实际根段数,<1 mm的根数为根尖数)、长度和鲜重,细根长度计算采用交叉法测量(Bohn, 1985),然后结合土壤体积换算成根长密度。
3 结果分析 3.1 植物篱复合经营系统根系的数量分布格局大于1 mm粗根的数量分布如图 2所示。可以看出,在0~45 cm土层,新银合欢粗根主要分布在植物篱带及两侧0.5 m范围内,占总量的78%,在植物篱下侧作物带有少量分布;桑树粗根主要分布在植物篱带,占总数量的66%;脐橙根系主要分布在脐橙植株上侧该土层作物带,在植物篱带没有粗根分布。

|
图 2 植物篱复合经营系统植物篱、桑树和脐橙的≥1 mm粗根数量分布格局 Fig. 2 Distribution patterns of thick roots of leucaena, mulberry and orange(ϕ≥1 mm) 图例下同。 The same legend as below. |
45~70 cm土层这几种植物粗根的数量分布规律与0~45 cm相似,但是数量大大减少。植物篱和桑树粗根仍然主要分布在植物篱带,其余少量粗根在植物篱上下两侧作物带中呈对称分布,但数量远远低于脐橙,新银合欢粗根在植物篱带占55%,桑树粗根主要分布在植物篱带,占42%。该土层中新银合欢的粗根数量远远多于桑树和脐橙,且主要分布在植物篱带,说明新银合欢是直的深根系;脐橙在作物带的细根数量远远高于新银合欢和桑树;桑树根系主要沿植物篱带分布。
70~100 cm土层新银合欢和桑树的粗根数量较大,仍然主要分布在植物篱带,分别占总数量的44%和29%,但是,在植物篱两侧作物带中的分布偏向于植物篱上侧的作物带,特别是新银合欢向作物带深层土壤扩展比较大。
植物篱复合经营系统小于1 mm根系的数量分布如图 3所示。可以看出,在0~45 cm土层,新银合欢细根主要分布在植物篱带面,占总量65%;在植物篱下侧作物带仅有少量分布;桑树根系分布在植物篱带及其下侧的土层,占56%;脐橙根系主要分布在该土层作物带,特别是脐橙植株上侧。

|
图 3 植物篱复合经营系统植物篱、桑树和脐橙的<1 mm细根数量分布格局 Fig. 3 Distribution of amount of fine roots of leucaela, mulberry and orange(ϕ < 1 mm) |
在45~70 cm土层,这3种植物根系数量分布规律与0~45 cm相似,但是数量大大减少。和桑树根系仍然主要分布在植物篱带,其余少量根系在植物篱上下两侧作物带中呈对称分布,但数量远远低于脐橙根系。该土层中新银合欢的粗根数量远远多于桑树和脐橙,且主要分布在植物篱带,这进一步说明新银合欢是直的深根系;桑树的根系主要沿植物篱带分布,占总根数的50%。
在70~100 cm土层新银合欢和桑树的细根数量较多,且主要分布在植物篱带,分别占总数量的48%和44%,其余的根系在植物篱两侧的作物带中分布,但偏向于植物篱上侧的作物带,特别是新银合欢,这说明植物篱根系向作物带深层土壤扩展。
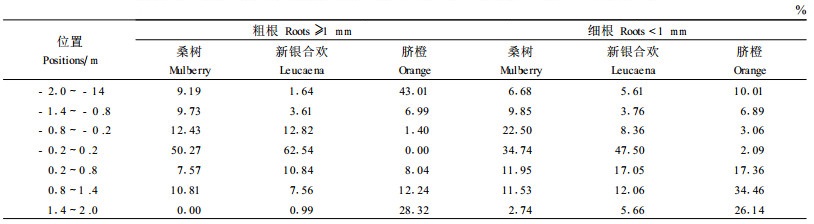
新银合欢、桑树和脐橙的粗根和细根数量在整个土壤剖面与植物篱的不同位置分布如表 1所示。新银合欢和桑树的粗根在植物篱带的分布占总数量的50%以上,在植物篱下侧0.8 m内有一些分布,脐橙粗根主要分布在距植株60 cm范围内,占71%;新银合欢细根在植物篱带及其相邻土壤中分布了近65%,桑树细根在植物篱带及其上下侧土壤分布了70%,脐橙细根在植物篱以外作物带土壤分布了97%以上。
|
|
大于1 mm根的根长密度分布如图 4所示。可以看出,在0~45 cm土层,新银合欢根长密度在植物篱带最大,其长度分别占总长度的79%;在植物篱下侧作物带有少量分布,桑树根系分布在植物篱带的土层占66%;脐橙根系主要分布在植株上侧作物带,占总长60%。

|
图 4 植物篱复合经营系统植物篱、桑树和脐橙的≥1 mm粗根根长密度分布格局 Fig. 4 Distribution of length density of thick roots of leucaena, mulberry and orange(ϕ≥1 mm) |
45~70 cm土层的根长密度分布规律与0~45 cm相似,但是根系数量大大减少。新银合欢和桑树根系仍然主要分布在植物篱带,分别占其总长的65%和35%,其余根系分布在植物篱上下两侧1 m以内作物带中,但下侧略高于上侧;脐橙根长密度在脐橙周围0.5 m范围最大,占总长度的78%。
70~100 cm土层新银合欢和桑树的粗根仍然主要分布在植物篱带,分别占总长度的63%和58 %,在植物篱两侧作物带中,新银合欢根系分布偏向于植物篱上侧的作物带,而桑树根系偏向于作物带下侧分布。脐橙在该土层基本上无粗根分布。
小于1 mm细根根长密度分布如图 5所示。可以看出,在0~45 cm土层,新银合欢细根主要分布在植物篱带,占总长度的49%,其余主要集中于在植物篱上侧作物带(20%);桑树细根在植物篱带占63%,其余主要集中在植物篱下侧作物带(21%);脐橙根系主要分布在植物篱带上侧1.5 m范围内(占细根总长79%)。

|
图 5 植物篱复合经营系统植物篱、桑树和脐橙的<1 mm细根长度分布格局 Fig. 5 Distribution of length of fine roots of leucaena, mulberry and orange trees(ϕ<1 mm) |
在45~70 cm土层,根长密度分布规律与0~45 cm相似,但是根系长度大大减少。新银合欢和桑树根系仍然主要分布在植物篱带(分别占其总长的38%和43%),其余少量根系分布在植物篱上下两侧作物带,新银合欢主要向上侧作物带分布(23%),桑树细根呈对称分布;脐橙细根主要分布于植株上侧0.75 m范围内,占细根总长63%。
70~100 cm土层新银合欢和桑树的细根仍主要分布在植物篱带,分布的细根占总细根长度的61%和71%,这说明新银合欢和桑树在深层土壤分布的直根系的特点;脐橙在该深度的细根主要分布于植株的下侧0.5 m范围内(66%)。
表 2可以看出,在整个土壤剖面,新银合欢及桑树粗根主要分布在植物篱带及其下侧(0.8 m以内),分布占总长的78%和68%,脐橙粗根在脐橙植株上下0.6 m范围内占了69%;在细根长度方面,新银合欢细根分布在植物篱及其上侧(68%),桑树细根集中分布在植物篱带及其下侧(79%),脐橙的细根分布不对称,向脐橙下侧分布到植物篱基部(占总长77%),而在脐橙植株上侧分布很少。
|
|

大于1 mm根系的生物量分布如图 6所示。可以看出,在0~45 cm土层,新银合欢和桑树粗根生物量集中在植物篱带,分别占粗根总生物量的98%和92%;脐橙粗根生物量主要集中于脐橙植株下侧距植株1.5 m范围内,占粗根总生物量66%。这说明用作植物篱的新银合欢和种植于植物篱中经济植物桑树的粗根生物量在作物带耕作层分布极少。

|
图 6 植物篱复合经营系统植物篱、桑树和脐橙≥1 mm粗根生物量分布格局 Fig. 6 Distribution of dry biomass of thick roots of leuceana, mulberry and orange(ϕ≥1mm) |
在45~70 cm土层,新银合欢和桑树的粗根生物量集中分布于植物篱带及其相邻0.5 m的两侧,其中植物篱带粗根生物量分别占总量的50%和58%;脐橙粗根生物量主要集中在距植株0.5 m范围内(75%)。
在70~100 cm土层,新银合欢和桑树的粗根生物量仍较大,主要分布在植物篱带,分别占粗根总生物量76%和64%;脐橙在各位置的粗根生物量均很小。
小于1 mm的细根生物量分布如图 7所示。可以看出,在0~45 cm土层,新银合欢细根生物量集中在植物篱带及其两侧,分别占细根总生物量的74%和83%,细根生物量相对粗根生物量要分散得多,并随与植物篱的距离增大而递减;脐橙细根生物量主要集中于植株下侧距植株1.5 m范围内,占细根总生物量75%。这说明新银合欢和桑树的细根在作物带耕作层分布极少。

|
图 7 植物篱复合经营系统植物篱、桑树和脐橙的<1 mm细根生物量分布格局 Fig. 7 Distribution of biomass of fine roots of leuceana, mulberry and orange (ϕ < 1 mm) |
在45~70 cm土层,新银合欢和桑树细根生物量集中在植物篱带及其上侧,分别占细根总生物量的64%和60%;脐橙细根生物量主要集中在植株及其下侧1.0 m范围内(61%)。
在70~100 cm土层新银合欢和桑树的细根数量较大,主要分布在植物篱带,在两侧作物带中的分布偏向于植物篱上侧的作物带;脐橙在该深度也有细根分布,生物量随与脐橙距离增大而递减。
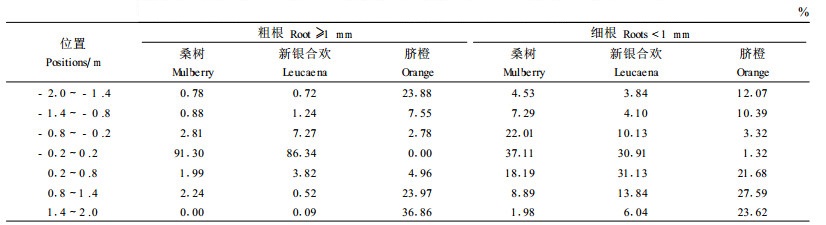
从表 3可以看出,新银合欢和桑树≥1 mm粗根干重在植物篱带的土壤中分别占了86%和91%;脐橙的粗根分布较散,在植株周围60 cm范围分布了84%,在植物篱带没有脐橙粗根。新银合欢的<1 mm细根生物量在植物篱及其上侧土壤中占了62%,桑树细根生物量在植物篱及其两侧土壤中占77%;脐橙细根在作物带土壤中分布了95%以上。
|
|
从以上分析可以看出,新银合欢和桑树≥1 mm粗根和<1 mm细根,不论数量、长度和生物量,都主要分布在植物篱带土壤中和植物篱临近两侧;脐橙的根系主要分布作物带中,在植物篱带的土壤中分布很少。本研究中植物篱间距采用4 m,在防治土壤侵蚀、土壤培肥和提高经济效益方面,效果比较理想(孙辉等,1999),许峰等(1999)在三峡的研究结果也表明植物篱最大间距应控制在5.9 m以内,才能达到植物篱有效控制水土流失、防止光竞争、提高土地利用率等目的。因此,在干旱河谷坡耕地上,等高固氮植物篱-桑树-脐橙模式中植物篱4 m间距对种植脐橙在经济方面和水土保持方面都是较合理的。
4 讨论对新银合欢等高固氮植物篱复合经营系统根系分布格局研究表明,植物篱根系主要集中分布在植物篱带及两侧0.5 m范围内,而且粗根在作物带中分布很少,在深土层主要集中在植物篱带,脐橙根系较浅,主要分布在脐橙植株1.0 m的范围内。这种分布格局使植物篱可以利用深层土壤的水分,吸收深层土壤的养分和表层淋溶的养分并通过刈割枝叶返还给作物带,与对农林复合经营系统中作物与木本植物关系的一些假设是相符的(Young, 1989),即通过调整农林复合经营各组分的时空配置来降低各组分间的养分竞争、促进和提高整个系统养分循环效率是可行的。
利用植物篱进行复合经营的主要目的在于坡耕地水土保持和土壤培肥,同时植物篱与农作物或经济作物尽量不发生或少发生水分、养分和光的竞争。水土保持可以通过植物篱的拦截过滤作用和形成生物梯地来实现(孙辉等,1999),已有的研究也表明植物篱在旱季利用土壤深层的水分,并在雨季增加水分向深层渗透,与农作物没有明显的水分竞争(孙辉等,2002b);在养分方面,植物篱系统中土壤养分不但沿土壤剖面深度分异,而且随与植物篱的距离不同而出现水平分异,二者之间根系在养分上没有明显的竞争。该系统中植物篱与脐橙的根系分布在各土壤层次均不重叠的根系分布格局,是该复合经营系统土壤水分和养分在坡面水平方向上分异的根本原因。这为像在干旱河谷这类水分时空分布极为不均的地区通过合理物种发展农林复合经营,充分利用不同深度土壤水分和养分资源,发展可持续旱地农林业提供了新的思路。
因为固氮植物篱通过细根周转,向作物带输入氮素和有机质;同时植物篱对坡面径流的拦截,沿植物篱根系下渗的水分总量和深度均比作物带大。这使植物篱土壤水分和养分状况都优于作物带,由于根系的向水性和向肥性,从而使桑树根系与植物篱根系在很大程度上是重叠分布的,桑树根系沿植物篱带分布,向作物带扩展很少。正因为如此,当地常规栽培的桑园和地埂桑每年需要施肥3~4次,而植物篱中种植的桑树不施肥也可生长良好,充分利用了植物篱带土地,提高经济效益(袁远亮等,2001;Tang et al., 2003)。
植物篱复合经营系统地下部分的研究不仅涉及植物篱和其他作物间的协同和竞争作用,而且在弄清该系统中各组分对水分和养分动态和利用模式、提高水分和养分利用效率和优化各组分的时空配置等方面也十分重要。Young(1989)认为,在农林复合经营中,地下部分在维持土壤肥力方面与地上部分同等重要,因为固氮植物篱通过细根周转,向较深层次的土壤输入氮素和有机质,但目前仍缺乏这方面的研究。因此,在固氮植物篱根系生物量和细根周转率对土壤养分动态的影响,还需进一步深入研究。
马秀玲, 陆光明, 徐祝龄, 等. 1997. 农林复合系统中林带与作物的根系分布特征. 中国农业大学学报, 2: 109-115. |
孙辉, 唐亚, 陈克明, 等. 1999. 固氮植物篱防治坡耕地土壤侵蚀效果研究. 水土保持通报, 19(6): 1-5. DOI:10.3969/j.issn.1000-288X.1999.06.001 |
孙辉, 唐亚, 王春明, 等. 2001. 等高固氮植物篱技术——山区坡耕地保护性开发利用的有效途径. 山地学报, 19(2): 125-129. DOI:10.3969/j.issn.1008-2786.2001.02.006 |
孙辉, 唐亚, 赵其国, 等. 2002a. 植物篱枝叶有机碳分解研究. 土壤学报, 39: 361-367. |
孙辉, 唐亚, 赵其国. 2002b. 干旱河谷区坡耕地植物篱种植系统土壤水分动态研究. 水土保持学报, 16(1): 84-87. |
孙辉, 唐亚, 赵其国, 等. 2003. 植物篱枝叶中P、K、Ca和Mg矿化过程研究. 应用与环境生物学报, 9: 1-6. DOI:10.3321/j.issn:1006-687X.2003.01.001 |
唐亚, 谢嘉穗, 陈克明, 等. 2001. 等高固氮植物篱技术在坡耕地可持续耕作中的应用. 水土保持研究, 8(1): 104-109. DOI:10.3969/j.issn.1005-3409.2001.01.023 |
许峰, 蔡强国. 1999. 等高植物篱在南方湿润山区坡地的应用:以三峡库区紫色土坡地为例. 山地学报, 17(3): 193-199. DOI:10.3969/j.issn.1008-2786.1999.03.001 |
袁远亮, 孙辉, 唐亚. 2000. 固氮植物篱梯埂套种桑树效益的初步研究. 中国生态农业学报, 8(2): 69-71. |
张小全, 吴可红. 2001. 森林细根生产和周转研究. 林业科学, 37(3): 126-138. DOI:10.3321/j.issn:1001-7488.2001.03.021 |
Agus F, Cassel D K. 1997. Soil-water and soil physical properties under contour hedgerow systems on sloping oxisols. Soil & Tillage Research, 40: 185-199. |
Bohn W. 1985.根系研究法.薛德榕译.北京: 科学出版社
|
Brunner I, Brodbeck S, Walthert L. 2002. Fine root chemistry, starch concentration, and 'vitality' of subalpine conifer forests in relation to soil pH. Forest Ecology and Management, 165: 75-84. DOI:10.1016/S0378-1127(01)00633-8 |
Imo M, Timmer V R. 2000. Vector competition analysis of a Leucaena-maize alley cropping system in western Kenya. Forest Ecology and Management, 126: 255-268. DOI:10.1016/S0378-1127(99)00091-2 |
Kang B T, Wilson G F, Sipkens L. 1981. Alley cropping maize(Zea mays L.) and leucaena(Leucaena leucocephala Lam de Wit) in southern Nigeria. Plant and Soil, 63: 165-179. DOI:10.1007/BF02374595 |
Lal R. 1989. Agroforestry systems and soil surface management of a tropical alfisol:Ⅰ. Soil moisture and crop yield. Agroforestry Systems, 8: 7-29. DOI:10.1007/BF00159066 |
Kang B T, Reynold L, Atta-Krah A N. 1990. Alley Farming. Advances in Agronomy, 43: 315-359. DOI:10.1016/S0065-2113(08)60481-2 |
Kiepe P. 1995. No runoff, no soil loss:soil and water conservation in hedgerow barrier system. Wageningen, Netherlands: Wageningen Agricultural University.
|
McIntyre B D, Riha S J, Ong C K. 1997. Competition for water in a hedge-intercrop system. Field Crops Research, 52: 151-160. DOI:10.1016/S0378-4290(96)01062-3 |
Lehmann J, Peter I, Steglich C, et al. 1998. Below-ground interaction in dryland agroforesrty. Forest Ecology and management, 111: 157-169. DOI:10.1016/S0378-1127(98)00322-3 |
Tang Ya. 2000. Contour hedgerow intercropping technology-its role and potential in sustainable management of sloping agricultural land in the Yangtze River basin. In: Proceedings of the Workshops on Capacity Building to Address Environmental Factors Contributing to Impacts of the Yantze River Flood Events. UNEP, UNCHS (Habitat), SEPA. Beijing: China Meteorological Press 235-240
|
Tang Ya, Zhang Y Z, Xie J S, et al. 2003. Incorporation of mulberry in contour hedgerows to increase overall benefits:a case study from Ningnan County, Sichuan Province, China. Agricultural Systems, 76: 775-785. DOI:10.1016/S0308-521X(02)00083-5 |
Young A. 1989. Ten hypotheses for soil-agroforestry research. Agroforestry Today, 1: 13-16. |