 2005, Vol. 41
2005, Vol. 41文章信息
- 方晰, 田大伦, 项文化, 闫文德, 康文星.
- Fang Xi, Tian Dalun, Xiang Wenhua, Yan Wende, Kang Wenxing.
- 杉木人工林林地土壤CO2释放量及其影响因子的研究
- Soil CO2 Release Rate and Its Effect Factors in Chinese Fir Plantation
- 林业科学, 2005, 41(2): 1-7.
- Scientia Silvae Sinicae, 2005, 41(2): 1-7.
-
文章历史
- 收稿日期:2004-05-18
-
作者相关文章


土壤CO2释放量既是土壤中微生物和植物根系进行生命活动的标志,也是碳循环的重要组成部分(陈全胜等,2003a;2003b)。测定土壤CO2释放量,确定其与环境因子的关系,不仅是评价生物学过程在生态系统碳收支平衡中作用的基础资料,而且对评测陆地生态系统在全球碳循环中的功能和地位也有着极其重要的意义。森林是陆地生态系统的主体,据估计,全球的碳有46.3%贮存在森林生态系统中,森林土壤的碳库占全球土壤总碳库的73%(Post et al., 1982),森林土壤CO2释放量往往被作为土壤微生物活性和土壤肥力乃至透气性的指标而受到重视(郑洪元等,1982;Neilson et al., 1990)。就森林生态系统而言,温度、水分和土壤营养条件等是影响土壤CO2产生和排放的重要且不易受控制的环境因素,了解林地土壤CO2释放量与环境因素之间的关系是建立估算区域森林生态系统CO2释放量各种模型的基础。同时,由温室效应和全球气候自然振荡引起的未来全球气温、降水量的任何变化均会在不同程度上影响土壤中许多微生物过程的速率,从而影响土壤CO2等温室气体的排放。近几十年来,在这方面人们开展了大量的研究工作,但大多数是在温带和热带地区进行的(Andrew et al., 1987;Ivan et al., 1993;Kucera et al., 1971;刘绍辉等,1998;Raich et al., 1992;孙向阳等,2001;刘建军等,2003;Toland et al., 1994;吴仲民等,1997;),在其他地方,特别是中国亚热带森林生态系统很少开展这项工作。杉木(Cunninghamia lanceolata)是我国南方亚热带地区特有的优良速生乡土用材树种。目前,我国杉木林面积达1 239.1万hm2,蓄积量为4.735 733亿m3,分别占全国人工林面积和蓄积的26.55%和46.89%。本研究探讨了杉木人工林地土壤CO2释放量与环境因子的关系,并建立杉木林地土壤CO2释放量与环境因子的相关模型,为进一步推动我国乃至全球气候变化的预测提供基础资料。
1 试验地概况试验地设于国家重点野外科学试验站、国家林业局重点森林生态系统定位观测站——中南林学院湖南会同森林生态系统定位研究站。地理坐标26°0′N,109°45′E,属典型的亚热带湿润性气候,年平均温度为16.8 ℃,年平均降水量在1 100~1 400 mm之间,年平均相对湿度为80%以上。海拔300~500 m,相对高度150 m以下,为低山丘陵地貌。土壤系震旦纪板溪系灰绿色板岩发育的山地黄壤,质地细,介于轻壤与粘壤之间,表土褐色至淡黄橙色,心土为橙黄色,非常适合杉木的生长。地带性植被为常绿阔叶林,以壳斗科(Fagaceae)的常绿树种栲属(Castanopsis)、青冈属(Cyclobalanopsis)、石栎属(Lithocarpus)为建群种,其次为樟科(Lauraceae)的樟属(Cinnamomum)、楠木属(Phoebe),山茶科(Theaceae)的木荷属(Schima)、山茶属(Camellia)以及木兰科(Magnoliaceae)、金缕梅科(Hamamelidaceae)、杜英科(Elaeocarpaceae)的一些树种组成。站内设有8个面积为2 hm2试验小集水区,相互平行且自然状况基本相似,平均坡度为25°,彼此相距不超过100 m。
本次试验在Ⅲ号集水区内进行。1987年,对该区1966年营造的杉木人工林皆伐后,当年炼山、整地,1988年以1 500~2 490株·hm-2营造杉木纯林。1988—1990年间,每年进行2次(5月、8月)全林抚育。现林分已郁闭,生长正常,郁闭度0.8~0.9,林内有少量的杜茎山(Maesa japonica)、木姜子(Litsea cubeba)、油桐(Fordii hemsrasiticus)和冬青(Ilex chinensis)等灌木,以及华南毛蕨(Cyclosorus parasiticus)、狗脊蕨(Woodwardia japonica)、铁芒箕(Dicranopteris dichotoma)、地菍(Melastoma dodecandrum)等草本植物。
2 研究方法 2.1 实验方法根据林地的地形、地势等特点,在Ⅲ号集水区内,在海拔330 m的山麓和350 m的山坡中上部分别定点测定和采样,于1996年3月底到11月底,每月中旬测定3天(晴天、阴天、雨天各1天),每天每个点重复采样2~3个,取其平均值作为当天的测定值,再以3天平均值作为该月份的测定值进行统计分析。
林地土壤CO2释放量的测定:采用田间容量法即用碱液Ba(OH)2吸收,HCl滴定,也称碱液静态吸收法(郑洪元等,1982)。具体操作如下:将瓶口直径为6 cm,容积为1 L的广口瓶嵌入土壤表面5 cm,瓶口周围压紧,瓶口事先用干净纸包住,以免使瓶口弄脏,放置30 min或更长时间后迅速取出,用橡皮塞塞紧(橡皮塞上有一小孔,用另小橡皮紧塞),从橡皮塞上小孔加入20 mL 0.025 mol·L-1 Ba(OH)2溶液,振荡5 min,用0.05 mol·L-1 HCl滴定,以酚酞指示剂。与此同时,另取一个同样大小的广口瓶放置于空气中,暴露同等的时间,作为对照。根据以下公式计算出林地土壤CO2(g·m-2h-1)释放量(R):
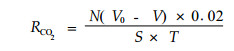
|
式中:N为盐酸(HCl)浓度(mol·L-1),V0为对照试样(空白试样)所消耗盐酸体积数(mL),V为试样所消耗盐酸体积数(mL),S为瓶口与土壤接触的表面面积(m2),T为采样时间(h)。
在测定林地土壤CO2释放量的同时,在林内的气象常规观测站内,观测林内1.5 m处空气温度;采用曲管地温表同点观测土壤表层5、10、15、20 cm处的土壤温度,采0、10、20 cm处的土壤测定其自然含水量。采用质量法测定土壤的自然含水量(中国土壤学会农业化学专业委员会,1984),计算公式为:

|
每一个月份分4个层次(0~15、15~30、30~45、45~60 cm)重复2~3次采取林地土壤样品测定土壤有机碳含量和全N含量,取其平均值作为每一个月的结果进行统计分析。
土壤样品中有机碳素含量采用重铬酸钾—水合加热法测定,全N用半微量凯氏法(中国土壤学会农业化学专业委员会,1984)测定。
2.2 数据处理试验数据采用EXCEL、SPSS10.0统计软件处理。
3 结果 3.1 林地土壤CO2释放量、土壤温度、土壤含水量、土壤C/N比和林内气温的季节动态由表 1可以看出,杉木人工林林地土壤CO2释放量变化范围为0.100 5~0.559 0 g·m-2h-1,变异系数为36%。土壤表层5、10、15、20 cm及林内的温度(℃)分别在8.2~29.4、10.0~24.6、10.7~24.4、11.6~24.5、12.1~24.5、7.2~34.1之间变动,变异系数为23%~29%。土壤0、10、20 cm的含水量(%)分别为26.73%~32.00%、23.00%~28.85%、22.00%~29.44%,变异系数为5.76%~3.47%。土壤0~15、0~45的C/N比分别为10.00~15.84、8.90~11.65,变异系数为8.47%~15.5%。
|
|
由图 1可以看出,在整个试验期间(4—11月份),杉木人工林林地土壤CO2释放量季节变化呈单峰曲线,总体趋势是夏季高,其他季节低,与对应的土壤温度、水分、C/N比、林内温度的季节变化基本一致,即随当地雨季的到来,林内气温和土壤温度上升、水分增加及C/N比的提高,CO2释放量表现为上升趋势,在夏季(8月份)出现最大值,达0.559 0 g·m-2h-1;随着当地雨季的结束,林内气温和土壤温度下降,水分减少,土壤C/N比下降,林地土壤CO2释放量表现为下降趋势,冬季(11月份)出现最低值,为0.105 2 g·m-2h-1。这说明雨季是植物生长旺盛时期,植物根系呼吸强度大,土壤微生物活性增强。

|
图 1 林地土壤CO2释放量、温度、土壤含水量、土壤C/N比季节动态 Fig. 1 Seasonal dynamics of soil CO2 release rate, temperature, soil water content and soil C/N ratio |
整个试验期间,林地土壤CO2释放量和土壤各层(0、5、10、15、20 cm)温度有着共同的变化趋势(见图 1)。林地土壤CO2释放量与土壤各层(0、5、10、15、20 cm)温度的相关性分析结果(图 2)表明,林地土壤CO2释放量与土壤表层或亚表层的温度均存在显著的线性正相关关系(R2>0.643 7)。在所有土壤温度指标与土壤CO2释放量之间的关系中,地表温度与土壤CO2释放量的关系最密切,其次是10 cm处的温度。根据图 2的相关方程可计算出,当土壤表层温度由10 ℃上升到20 ℃时,杉木林地土壤CO2释放量增加280%(即Q10=2.80),由20 ℃上升到30 ℃时,Q10=1.64,平均Q10=2.22。同样,可计算出土壤5、10、15、20 cm处的平均Q10分别为1.94、2.28、2.51、2.23。

|
图 2 林地土壤CO2释放量与土壤温度的线性关系 Fig. 2 The relationships between soil CO2 release rates and soil temperature |
从图 1可以看出,林地土壤CO2释放量随土壤含水量增加而增大,说明林地土壤CO2释放量与土壤含水量也有一定的相关性,这可能是由于土壤含水量对土壤呼吸强度的影响。统计分析结果(图 3)表明,已郁闭杉木人工林林地土壤CO2释放量与土壤各层(0、10、20 cm)含水量有较明显线性正相关(R2≥0.256 8),其中与土壤表层含水量的相关性最高。

|
图 3 林地土壤CO2释放量与土壤含水量的线性关系 Fig. 3 The relationships between soil CO2 release rates and soil water content |
林地土壤CO2释放量与土壤C/N比的相关性分析结果(见图 4)表明,林地土壤CO2释放量与土壤C/N比存在一元线性正相关关系(R2≥0.370 8)。

|
图 4 林地土壤CO2释放量与土壤中C/N比的关系 Fig. 4 Relationship between soil CO2 release rates and soil C/N ratios in stands |
如图 1所示,杉木林地土壤CO2释放量与林内气温的季节变化基本上是同步的,但是两者的动态曲线并非完全吻合,说明在这一过程中林内气温不是唯一的影响因子,在<26 ℃时,两者的相关性非常高。对林地土壤CO2释放量与林内气温连续1年24次同步实测数据的相关性分析结果(图 5)表明,林地土壤CO2释放量与林内气温之间存在显著的线性正相关关系(R2=0.838 5)。当林内气温由10 ℃上升到20 ℃时,杉木林地土壤CO2释放量增加254%(即Q10=2.54);由20 ℃上升到30 ℃时,Q10=1.61,平均Q10=2.08。可见,在一定温度范围内,林内气温每上升10 ℃,杉木林地土壤CO2释放量增加208%。

|
图 5 林地土壤CO2释放量与林内气温的关系 Fig. 5 Relationship between soil CO2 release rates and air temperature in stands |
为了进一步探明环境因子对杉木人工林林地土壤CO2释放量的综合影响,在这里采用Pearson偏相关方法分析杉木人工林林地土壤CO2释放量与土壤各层(0、5、10、15、20 cm)温度、土壤各层(0、10、20 cm)含水量、土壤(0~15、0~45 cm)C/N比、林内气温等环境因子的相互关系。结果表明,郁闭杉木人工林林地土壤CO2释放量与林内气温相关性最大,偏相关系数达0.916(p<0.01),这是郁闭杉木人工林林地土壤CO2释放量呈现出明显的季节变化节律的原因;与土壤各层(0、5、10、15、20 cm)温度的偏相关系数均在0.818以上(p<0.01);与土壤C/N比和土壤各层(0、10、20 cm)含水量的偏相关系数为0.452~0.612(p>0.05)。从t检验的p值来看,杉木林林地土壤CO2释放量与林内气温、土壤各层(0、5、10、15、20 cm)温度的偏相关达到了极显著水平(p<0.01),而与土壤C/N比、土壤各层(0、10、20 cm)含水量的偏相关性不显著(p>0.05)。表明杉木人工林林地土壤CO2释放量主要受到林内气温和土壤各层(0、5、10、15、20 cm)温度的影响,且林地土壤CO2释放量与各环境因子的关系有一定的差异。
鉴于Pearson偏相关方法分析的结果,以土壤各层(0、5、10、15、20 cm)温度、土壤各层(0、10、20 cm)含水量、土壤(0~15、0~45 cm)C/N比、林内气温为变量,采用多元线性回归中的Stepwise方法进行逐步回归,分析结果(表 2)表明,杉木林地土壤CO2释放量(y)与林内气温(x1)的关系模型为:
|
|

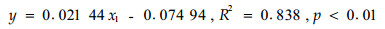
|
(1) |
杉木林地土壤CO2释放量(y)与林内气温(x1)、土壤10 cm处含水量(x2)的关系模型为:
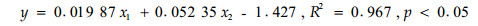
|
(2) |
F检验表明,方程(1)、(2)回归均为显著。但经各个自变量的偏回归系数检验表明,土壤10 cm处含水量的偏回归不显著,即土壤10 cm处含水量在方程(2)中不起明显的作用。
4 结论与讨论湖南省会同杉木中心产区内,已郁闭杉木人工林林地土壤CO2释放量一般在0.105 2~0.554 4 g·m-2h-1之间变化。土壤中的CO2,主要来源于土壤微生物、植物根系、土壤动物和原生动物的生命活动,这些生命活动均受到气温、土壤温度、水分、营养状况等因素的影响。作为土壤生物总体代谢强度反映的土壤CO2释放量,一年中随着土壤温度、湿度、C/N比、林内气温等外界环境因子的季节变化必然有较强的季节变化。
已郁闭杉木人工林林地土壤CO2释放量与林内气温、土壤各层的温度在一定范围内呈一元线性正相关。温度对植物根系的呼吸、土壤微生物的活性以及有机物质的分解等均有促进作用,因而对土壤释放CO2的影响是显著的。有研究认为,夏季土壤CO2释放强度是冬季的5倍(杜宝华,1994),本研究结果与这一结论接近。许多学者采用Q10值来说明土壤CO2释放量随着温度的变化。温带地区,当土壤温度每上升10 ℃时,辽东栎(Quercus liaotungensis)林地土壤释放CO2的强度增加151%(蒋高明等,1997)。中亚热带东部,在-5~35 ℃地表温度范围内,当地面温度上升10 ℃时,青冈(Cyclobalanopsis glauca)常绿阔叶林地土壤呼吸增加255%,毛竹林地增加211%,茶园地增加175%(黄承才等,1999)。本研究中,在一定土壤温度范围内,土壤表层温度每上升10 ℃,杉木林地土壤CO2释放量增加222%,高于农业土壤的Q10=2.00(杜宝华等,1985),即在一定的温度范围内有利于植物根系的呼吸、土壤微生物的活性,而土壤温度提高能改变土壤有机质的物理化学状态,促使一部分有机质更为有效。
已郁闭杉木人工林林地土壤CO2释放量与土壤各层含水量呈一元线性正相关。一般认为,合适的土壤湿度(最大田间持水力)有利于可溶性有机质的淋溶分解,土壤通透性增加,促进微生物、植物的生长和活动,当土壤处于过干或过湿状态时,土壤呼吸会受到抑制(布雷迪,1982;陈全胜等,2003)。据报道(蒋高明等,1997),湿度越高,意味着土壤越湿润,CO2在土壤中的溶解随之加快,从而提高土壤中CO2浓度,土壤释放CO2的强度也因之提高,湿润土壤与干燥土壤CO2释放量分别为17.8和11.8 g·m-2h-1。
已郁闭杉木人工林林地土壤CO2释放量与土壤C/N比也呈一元线性正相关。一般认为土壤有机碳的保持在很大程度上取决于土壤全N的含量,即当土壤C/N比高时,普通腐生微生物(如细菌、真菌等)活性提高、繁殖快,产生大量的CO2,硝化细菌或多或少地处于抑制状态;当土壤C/N比低时,硝化作用旺盛,土壤中的硝酸盐多,产生CO2较少,且这两种状态维持时间的长短,土壤C/N比是一个重要的因子(布雷迪,1982)。因此,可考虑在不影响林木生长的条件下,通过人为宏观调控林内气温、土壤温度、土壤湿度及土壤中的C/N比等环境因子改变土壤作为温室气体源的强度,以改善大气环境质量。
相关分析表明,林地土壤CO2释放量与林内气温相关性最大且为极显著相关,偏相关系数达0.916(p<0.01),这是已郁闭杉木人工林林地土壤CO2释放量呈现明显季节变化的主要原因;与土壤各层(0、5、10、15、20 cm)的温度也达到极显著相关,偏相关系数均在0.818(p<0.01)以上,这表明林内气温和土壤各层(0、5、10、15、20 cm)温度是影响杉木人工林林地土壤CO2释放量的主要因子。
值得注意的是,本研究的相关分析表明,土壤各层(0、10、20 cm)含水量与林地土壤CO2释放量的相关关系不显著(p>0.05)。这是因为,在自然条件下土壤水分对土壤呼吸的影响主要是通过对植物和微生物的生理活动、微生物的能量供应和体内再分配、土壤的通透性和气体的扩散等的调节和控制实现的(陈全胜等,2003a; 2003b)。二者的关系非常复杂,并非一种简单的因果关系。有研究表明,在土壤水分含量充足、不成为限制因子的湿润地区,土壤呼吸与温度成正相关关系,但在土壤水分含量成为限制因子的干旱、半干旱地区,土壤水分含量与土壤温度对土壤呼吸共同起作用(Nakane et al., 1985;Mathes et al., 1985;刘绍辉等,1997;黄承才等,1999)。本研究区属于典型的亚热带湿润气候区,年降水量约为1 100~1 400 mm,同时各个自变量的偏回归系数检验表明,土壤10 cm处含水量的偏回归系数不显著,即可初步确定该林区的土壤水分不成为杉木林地土壤CO2释放量限制因子,杉木林地土壤CO2释放量的变化主要受到林内气温的影响。同时,方程(1)的预测值与实测值相拟较好,因而在正常的范围内,可根据方程(1)结合林内气温的定位观测来预测杉木人工林林地土壤CO2释放量动态。
布雷迪NC(美)著.1982.南京农学院土化系等译.土壤的本质与性状.北京: 科学出版社
|
陈全胜, 李凌浩, 韩兴国, 等. 2003a. 水分对土壤呼吸的影响及机理. 生态学报, 23(5): 972-978. |
陈全胜, 李凌浩, 韩兴国, 等. 2003b. 水热条件对锡林河流域典型草原退化群落土壤呼吸的影响. 植物生态学报, 27(2): 202-209. |
杜宝华. 1994. 作物冠层内二氧化碳源、流研究现状与展望. 中国农业气象, 15: 44-48. |
杜宝华, 洪佳华, 司可君, 等. 1985. 农田土壤CO2释放量的测定. 农业气象, (2): 63-67. |
黄承才, 葛滢, 常杰, 等. 1999. 中亚热带东部三种主要木本群落土壤呼吸的研究. 生态学报, 19(3): 324-328. DOI:10.3321/j.issn:1000-0933.1999.03.006 |
刘建军, 王得祥, 徐德应, 等. 2003. 秦岭天然油松、锐齿栎林地土壤呼吸与CO2释放. 林业科学, 39(2): 8-13. |
刘绍辉, 方精云. 1998. 北京山地温带森林的土壤呼吸. 植物生态学报, 22(2): 119-126. |
刘绍辉, 方精云. 1997. 土壤呼吸的影响因素及全球尺度下温度的影响. 生态学报, 17(5): 469-476. DOI:10.3321/j.issn:1000-0933.1997.05.003 |
蒋高明, 黄银晓. 1997. 北京山区辽东栎林土壤释放CO2的模拟实验研究. 生态学报, 17(5): 477-482. DOI:10.3321/j.issn:1000-0933.1997.05.004 |
孙向阳, 乔杰, 谭笑. 2001. 温带森林土壤中的CO2排放通量. 东北林业大学学报, 29(1): 34-39. DOI:10.3969/j.issn.1000-5382.2001.01.009 |
吴仲民, 曾庆波, 李意德, 等. 1997. 尖峰岭热带森林土壤C储量和CO2排放量的初步研究. 植物生态学报, 21(5): 416-423. DOI:10.3321/j.issn:1005-264X.1997.05.003 |
郑洪元, 张德生主编. 1992. 土壤动态生物化学研究法. 北京: 科学出版社.
|
中国土壤学会农业化学专业委员会. 1984. 土壤农业化学常规分析方法. 北京: 科学出版社.
|
Andrew M, Gordon Robert E, Schlentner Keith Van Cleve. 1987. Seasonal patterns of soil respiration and CO2 evolution following harvesting in the white spurce forest of interior Alaska. Can J For Res, (17): 304-310. |
Ivan J, Fernadez, Yowhan Son, et al. 1993. Soil Carbon Dioxide characteristics under different forest types and after harvest. Soil sci Soc Am J, 57: 1115-1121. DOI:10.2136/sssaj1993.03615995005700040039x |
Kucera C L, Kirkham D R. 1971. Soil respiration studies in tallgrass prairie in Missouri. Ecology, 52: 912-915. DOI:10.2307/1936043 |
Macfadyen A. 1970. Simple methods for measuring and maintaining the proportion of carbon dioxide in air, for use in ecological studies of soil respiration. Soil Biology and Biochemistry, 2: 9-8. DOI:10.1016/0038-0717(70)90020-9 |
Mathes K, Schriefer Th. 1985. Soil respiration during secondary succession influences of temperature and moisture. Soil Biol Biochem, 17(2): 205-211. DOI:10.1016/0038-0717(85)90116-6 |
Nakane K, Tsubota H, Yamamoto M. 1984. Cycling of soil carbon in a Japanese Red Pine Forest Ⅰ. Before a clear-felling. Bot Mag, 97: 39-60. DOI:10.1007/BF02488146 |
Neilson J W, Pepper I L. 1990. Soil respiration as an index of soil aeration. Soil Science Society of American Journal, 54: 428-432. DOI:10.2136/sssaj1990.03615995005400020022x |
Post W M, Emanuel W R, Paul J, et al. 1982. Soil carbon pools and world life zones. Nature, 298: 156-159. DOI:10.1038/298156a0 |
Raich J W, Schlesinger W H. 1992. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tullus, 44B: 81-99. |
Toland D E, Zak D R. 1994. Seasonal pattern of soil respiration in intact and clear-cut northern hardwood forest. Can J For Res, 24(8): 1171-1176. |