 2005, Vol. 41
2005, Vol. 41文章信息
- 张继义, 赵哈林, 崔建垣, 李玉霖, 苏永中.
- Zhang Jiyi, Zhao Halin, Cui Jianyuan, Li Yulin, Su Yongzhong.
- 沙地植被恢复过程中克隆植物分布及其对群落物种多样性的影响
- Effects of Clonal Plants on Community Structure and Function along a Restorational Gradient in Horqin Sandy Land
- 林业科学, 2005, 41(1): 5-9.
- Scientia Silvae Sinicae, 2005, 41(1): 5-9.
-
文章历史
- 收稿日期:2003-02-09
-
作者相关文章


2. 中国科学院寒区旱区环境与工程研究所 兰州 730000
2. Cold and Arid Regions Environmental and Engineering Research Institute, Chinese Academy of Sciences Lanzhou 730000
克隆植物是指在自然条件下能够进行无性繁殖以实现种群增长的一类植物(De kroon et al., 1997)。克隆植物在自然生态系统中广泛存在,并在许多生态系统中处于优势地位(Callagham et al., 1992; Prach et al., 1994)。由于克隆植物的重要性,近年来对克隆植物的研究受到了越来越多研究者的注意(宋明华等,2002)。研究证明,克隆植物具有较强的适应环境压力和抗扰动能力,在植物群落中起着重要作用(董鸣,1996;董鸣等,1999)。克隆植物具有很强的水平扩展能力,能有效地利用资源,迅速开拓新生境,扩大种群,形成单优势种的立地(Callagham, 1988; 董鸣,1996)。克隆植物在群落中的出现强烈地影响和制约着植物群落的空间格局与竞争关系,影响群落的物种多样性(De kroon et al., 1997)。
“密集型”(phalanx)和“游击型”(guerilla)被认为是克隆植物2种基本的克隆生长构型(Lovett, 1981; Schmid et al., 1985)。不同克隆生长构型的克隆植物在群落中出现的频率和分布的生境不同(De kroon et al., 1997; 宋明华等,2001)。克隆植物与非克隆植物对群落物种多样性的影响采取不同的机制,群落中克隆植物重要性同物种多样性之间的关系与植物的克隆生长构型及生境状况有关(宋明华等,2002)。
植被恢复是退化生态系统重建特别是荒漠化治理的重要途径和步骤。物种多样性是群落的重要特征,植被恢复过程物种多样性的变化反映植被的恢复程度,同时也是群落环境演变、种群侵入与扩散、竞争作用等生态过程共同作用的结果(Richard et al., 1987)。本文以科尔沁沙地草地植被不同恢复程度的群落类型为对象,研究了克隆植物在恢复梯度上的分布及其对群落物种多样性的影响,以利于深入认识克隆植物的生态作用及其对植被恢复和群落演替的意义,为该地区的植被恢复和退化沙地生态系统重建提供科学依据。
1 材料与方法 1.1 自然概况研究在位于科尔沁沙地中南部的内蒙古自治区奈曼旗境内中国科学院奈曼沙漠化研究站进行,地理位置120°41′E, 42°54′N。该地区属半干旱气候,年平均降水量366 mm,年蒸发量1 935 mm,年均气温6.5 ℃, 1月平均气温-12.7 ℃,7月平均气温23.7 ℃,≥10 ℃积温3 000 ℃以上,无霜期150 d。年均风速3.5 m·s-1,8级以上大风年均21次,主要发生在春季3—5月。土壤类型为沙质栗钙土,经破坏后则退化为流动风沙土。沙土基质分布广泛,风沙活动强烈(赵哈林等,1998)。
在科尔沁沙地,一方面由于沙土基质干燥疏松的不稳定性和干旱多风等气候条件的耦合,使得该地区在过度放牧和过度开垦等人为不合理利用的情况下,植被极易破坏,土地沙化,成为裸露流动沙丘。另一方面,由于沙土良好的水分下渗性能和保水能力,流沙上常具有较高的土壤含水量,为植被恢复提供了良好的水分条件。在大量天然种源存在的情况下和禁止放牧、禁止开垦等良好的保护措施下,存在着植被快速建立并向稳定植被类型恢复的可能,这也为植被恢复演替系列上的群落调查取样提供了可能。
由流动沙丘到半固定沙丘再到固定沙丘,每一阶段都发育着典型的群落类型。以流动沙丘上先锋植物——沙米群落为植被恢复的起点,植被恢复的过程也就是松散沙粒基质的流动性不断降低,沙地逐渐得到固定的过程。流动沙丘上部建立的植物群落几乎毫无例外地以沙米为优势种,盖度小,为10%~17%,群落的斑块化程度和空白斑块比例较高,随后逐渐向沙米-狗尾草和狗尾草-沙米,以及五星蒿-狗尾草群落演变,并伴随差巴嘎蒿的侵入和扩展。半固定沙丘则以差巴嘎蒿为优势种,群落盖度20%~30%。而在固定沙地,随着植被发育和沙地固定程度的提高,差巴嘎蒿种群衰退,猪毛菜、三芒草、狗尾草等成为优势种,并有胡枝子、扁蓿豆、隐子草等多年生植物的侵入和扩展,群落盖度30%~40%。随着沙地固定时间的延长,植被发育程度进一步提高,群落演变为以隐子草等多年生植物为优势种,群落盖度进一步提高,为40%~90%。沙地环境的变化与群落类型的演替变化相一致和相对应(李胜功等,1997),因此把沙地环境从流动沙丘、到半固定沙丘、再到固定沙丘的几个明显的阶段作为植被恢复程度和群落演替阶段的划分。
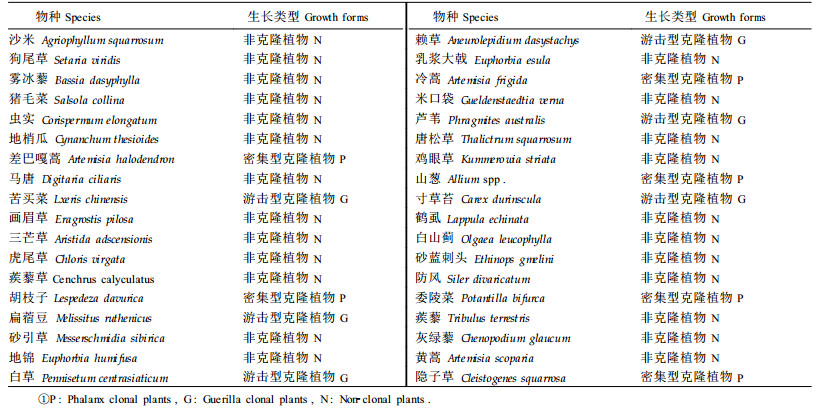
1.2 调查取样用时-空替代的方法,选取围封禁牧的分别处于流动沙丘、半固定沙丘和固定沙丘等不同恢复阶段和恢复时间分别为1、3、5、12、20和30年的6个典型群落样地进行调查。每群落类型样地上取互相平行的3条样带,样带长度100 m,样带间距50 m,样带上每隔10 m取样方1个,样方面积按照草本植被常规调查方法取1 m2。每群落类型视群落斑块面积大小取样方10~45个。样方中调查各植物种类的密度、高度和盖度。观察记录样地内植物的形态特征;对有根茎的物种,挖出其地下部分,判定植物类型(克隆植物,非克隆植物)及克隆植物生长构型(密集型,游击型)(宋明华等,2002)。恢复梯度上出现的植物种类及其生长类型见表 1。
|
|
相对盖度(%)=种i的盖度/样方内所有种的盖度和;
相对高度(%)=种i的高度/样方内所有种的高度和;
相对密度(%)=种i的密度/样方内所有种的密度和;
重要值Ii=(种i的相对盖度+种i的相对高度+种i的相对密度)/300;
多样性指数用Shannon-Wiener指数表示:Hiv=-∑Ii*ln(Ii)。
2 结果与分析 2.1 恢复梯度上物种丰富度的变化如图 1A所示, 克隆植物物种丰富度随着恢复时间延长而增加, 在固定沙地阶段达到最高, 以后并不随恢复时间的延长而增加。非克隆植物的物种丰富度总体上随着恢复而增加,但在半固定沙地的差巴嘎蒿阶段成下降趋势,以后又随着演替进入固定沙地阶段而增加。在整个恢复过程中,非克隆植物的物种丰富度都远远超过克隆植物。密集型克隆植物与游击型克隆植物的物种丰富度随恢复梯度的变化趋势一致(图 1B),且二者基本相同。

|
图 1 恢复梯度上克隆植物物种丰富度的变化 Fig. 1 Variation of species richness of clonal plants in the process of restoration |
克隆植物的重要值在恢复阶段的前期随恢复时间延长而增加,到恢复时间为12年的半固定沙地克隆植物的重要值达到一个顶峰,之后下降到一个中等水平(20年时),并随着恢复程度提高继续增加(30年时)(图 2A)。而非克隆植物的重要值在恢复的开始阶段(1~5年)远远高于克隆植物,到恢复的中期阶段(12年时)克隆植物的重要值远远超过了非克隆植物,二者的对比关系与前期相比出现了一个逆转。之后,非克隆植物的重要值再次超过了克隆植物,但二者的差距比恢复前期缩小。密集型克隆植物的重要值在恢复梯度上的变化与克隆植物的变化完全类似(图 2B),在恢复前期至中期,密集型克隆植物的重要值随恢复显著增加,之后下降到一个中等水平,到30年时又有所增加。游击型克隆植物的重要值在恢复阶段上呈缓慢增加,到恢复的较高阶段(30年时)明显下降。恢复梯度上密集型克隆植物的重要值始终超过游击型克隆植物,而且在恢复的前期二者的差距较小,在中期(12年时)差距最大,随后差距又有所缩小,但仍超过前期。

|
图 2 恢复梯度上克隆植物重要值的变化 Fig. 2 Variation of the importance value of clonal plants in the process of restoration |
在群落恢复的早期(1年和3年),克隆植物的数量非常少(图 3A,B),对群落物种多样性的影响也很小;在恢复期为5年的群落中,Shannon-Wiener指数与群落中克隆植物的重要值呈相关微弱的抛物线型(图 3C);在恢复期为12年的半固定沙地差巴嘎蒿群落中,Shannon-Wiener指数与克隆植物的重要值呈明显的半抛物线型(图 3D);恢复期为20年时,群落的物种多样性指数与克隆植物重要值为全抛物线型(图 3E);在恢复期为30年的固定沙地隐子草群落中,Shannon-Wiener指数与克隆植物重要值呈线型负相关(图 3F)。

|
图 3 恢复梯度上克隆植物重要值与物种多样性的关系 Fig. 3 The relationships of importance value of clonal plants and species diversity in the process of restoration A~F分别为恢复时间1、2、5、12、20和30年. A to F is the restoration time 1 a, 3 a, 5 a, 12 a, 20 a and 30 a respectively. |
沙地植被恢复过程中存在着明显的物种更替、群落环境变化等次生演替现象。群落演替的实质是物种的更替,是物种的环境适应性、群落环境的演变、种间竞争作用等生态因素共同作用的结果(Richard et al., 1987)。在科尔沁沙地从流动沙地开始的植被恢复过程中,随着植被的发育,群落环境发生变化。流动沙地上植被少,对土壤水分的消耗少,同时沙质土壤的疏松性质能对天然降水充分地纳蓄和下渗,因而流动沙地上水分条件好。但由于缺乏植被保护,风沙活动强烈,风蚀严重,基质条件极不稳定,不利于植物繁殖体的传播和定居。随着恢复的进行和植被的发育,群落盖度提高,生态功能增强,基质条件趋向稳定,但植被的发育加剧了土壤水分消耗,土壤环境旱化倾向不断加剧和发展。同时植被发育伴随成土过程的发展和有机质的积累,土壤肥力提高。在整个恢复梯度上,克隆植物、非克隆植物的物种丰富度不断增加,这是群落总体环境改善的结果。恢复后期(20和30年时)群落中物种丰富度的变化不大,说明群落物种丰富度已接近于区域物种库的饱和值。可以预言,随着演替的进一步发展,群落物种丰富度将增加缓慢。
在群落环境变化和物种更替的过程中,克隆植物的生态功能占据着重要的位置。从表面上看,在恢复过程中非克隆植物在群落中的重要值总体上超过了克隆植物,只有半固定沙地的差巴嘎蒿群落阶段克隆植物的重要值显著超过了非克隆植物。但是当把这种情况与时间尺度联系起来分析后就可以看到,克隆植物的生态功能实际上在群落中起主导作用。在整个恢复阶段上群落结构和功能的变化可以分为两个阶段,第1个阶段(1~12 a)是差巴嘎蒿种群的增长阶段。第2个阶段(20~30 a)是差巴嘎蒿种群的衰退阶段和另一种优势克隆植物隐子草的增长阶段。差巴嘎蒿是一种密集型克隆植物,具有沙生适应特性,耐沙埋,枝条细而柔软,沙埋后能产生不定根,并萌发大量枝条。经过一个较长时间后,差巴嘎蒿便能积累很高的地上生物量。1~5年是差巴嘎蒿的侵入、定居过程,因而这一时期克隆植物在群落中的重要值并不明显。恢复至12年时差巴嘎蒿已增长为群落的优势种群,这时群落中克隆植物与非克隆植物重要值的对比出现了逆转。差巴嘎蒿群落较高的地上生物量促使群落环境进一步改善:较高的地上生物量导致高的水分消耗,土壤旱化进一步发展;稠密的枝叶对地表的保护作用加强,阻止风蚀,拦截降尘,细土物质沉积,土壤结皮形成。群落环境从半固定沙地转向固定沙地。由于群落环境的改变,差巴嘎蒿失去了原来使其种群迅速增长的沙埋条件,土壤旱化也增强了一些旱生杂类草植物如猪毛菜、三芒草的竞争能力,最终差巴嘎蒿种群在新的环境下开始衰退。恢复至20年时沙地环境进入固定沙地的初期,差巴嘎蒿种群的衰退使克隆植物在群落中的重要值降低。这时另一种克隆植物隐子草已侵入群落并开始种群的增长过程,到恢复时间为30年时由于隐子草种群的增长(其在群落中的重要值上升至24.028%)使克隆植物在群落中的作用上升。隐子草是一种极耐旱的密集型克隆植物,具有发达的根系,其根系细而致密,主要分布在10~40 cm的土层范围内,能够主要以吸收天然降水维持生存并保持种群稳定。因而,隐子草能够成为固定沙地的建群种。可以预测,随着演替的进一步发展,以隐子草为主的克隆植物在群落中的重要值将再一次超过非克隆植物。
在整个恢复梯度上,密集型克隆植物的重要值始终明显超过游击型克隆植物,游击型克隆植物的重要值始终处于一个较低的水平(宋明华等,2002)。
克隆植物对群落物种多样性的影响在整个恢复梯度上有2个明显的时期。一是在恢复时间为12年的半固定沙地时期,由于克隆植物对空间的充分占据,使群落物种多样性下降;二是在固定沙地的初期(恢复时间20年时),由于密集型克隆植物差巴嘎蒿种群的衰退为其它物种的侵入提供了空间,群落的物种多样性增加。当植被盖度较高时,空间的利用充分,种间竞争较强,密集型克隆植物在群落中容易形成单优势种的立地,其对群落生态功能的维持起重要作用。但是过高的优势度所导致的竞争排除作用将在小尺度(样方尺度)上抑制其它物种侵入,从而导致群落物种丰富度和物种多样性的下降(宋明华等,2002)。
董鸣. 1996. 植物的克隆生长及其与资源异质性的关系:觅食行为. 植物学报, 38(10): 828-835. |
董鸣, 阿拉腾宝, 邢雪荣, 等. 1999. 根茎禾草披针叶黄花的遗传特征和分株种群特征. 植物生态学报, 23(4): 302-310. DOI:10.3321/j.issn:1005-264X.1999.04.002 |
宋明华, 董鸣, 蒋高明, 等. 2001. 沿东北样带上克隆植物重要性与环境条件的关系. 生态学报, 21(7): 1095-1103. DOI:10.3321/j.issn:1000-0933.2001.07.009 |
宋明华, 陈玉福, 董鸣. 2002. 鄂尔多斯高原风蚀沙化梁地克隆植物的分布及其与物种多样性的关系. 植物生态学报, 26(4): 396-402. DOI:10.3321/j.issn:1005-264X.2002.04.003 |
赵哈林, 刘新民, 李胜功, 等. 1998. 科尔沁沙地脆弱生态环境的基本属性特征和成因分析. 中国沙漠, 18(Supp.2): 10-16. |
李胜功, 赵爱芬, 常学礼. 1997. 科尔沁沙地植被演替的几个问题. 中国沙漠, 17(Supp.1): 25-32. |
Callagham T V.1988. Physiological and demographic implications of modular const ruction in cold environments. In: Davy A J, Hutchings M J & Watkinson A R eds. Plant population ecology. Oxford: Blackwell Scientific Publications, 113-135
|
Callagham T V, Carlsson B A, Jónsdóttir I S, et al. 1992. Clonal plants and environmental change: introduction to the proceedings and summary. Oikos, 63: 341-347. DOI:10.2307/3544959 |
De kroon H, Van groendael J.1997. The ecology and evolution of clonal plants. Leiden, The Netherlands: Backhuys Publishers
|
Lovett D L. 1981. Population dynamics and local specialization in a clonal plant Ranunculus repens. I. The dynamics of ramets in contrasting habitats. Journal of Ecology, 69: 743-755. DOI:10.2307/2259633 |
Prach K, Pysěk P. 1994. Clonal plants—what is their role in succession. Folia Geobetanica Phytotaxonomica, 29: 307-320. DOI:10.1007/BF02803803 |
Richard S I, Nancy J H, Tilman D, et al. 1987. Old-field succession on a Minnesota sand plain. Ecology, 68(1): 12-26. DOI:10.2307/1938801 |
Schmid B, Harper J L. 1985. Clonal growth in grassland perennials Ⅰ. Desity and pattern dependent competition between plants with different growth forms. Journal of Ecology, 73: 793-808. |