 2004, Vol. 40
2004, Vol. 40文章信息
- 程水源, 王燕, 李俊凯, 费永俊, 朱桂才.
- Cheng Shuiyuan, Wang Yan, Li Junkai, Fei Yongjun, Zhu Guicai.
- 内源激素含量与银杏叶中类黄酮含量的关系
- Study on the Relationship between the Endogenous Hormones and Flavonoids in Ginkgo biloba Leaf
- 林业科学, 2004, 40(6): 45-49.
- Scientia Silvae Sinicae, 2004, 40(6): 45-49.
-
文章历史
- 收稿日期:2003-01-22
-
作者相关文章


银杏叶片中类黄酮具有重要的药用价值而成为近年来研究的焦点。在类黄酮的合成代谢过程中,内源激素的研究有许多报道,较为肯定的说法有:乙烯(ethylene, ETH)、脱落酸(abscisic acid, ABA)促进类黄酮包括花青苷形成,而赤霉素(gibberellin, GA)效果相反,生长素(3-indole acetic acid, IAA)暂无定论,细胞分裂素(cytokinin, CTKs)报道较少,也没有取得较为一致的结论(程水源等,2000;Max, 1990)。关于内源激素的作用机理大多与促进成熟和有关酶活性有关(Max, 1990),但至今为止,这方面的研究在银杏(Ginkgo biloba)叶上仍是空白。本试验以银杏叶为试材,探讨内源激素与类黄酮的关系,目的在于明确内源激素在银杏叶中类黄酮形成中所起的作用与地位,为调控银杏叶中类黄酮合成提供理论与实践资料。
1 材料与方法 1.1 选材、取样及预处理试材选于湖北农学院银杏园,品种为7年生“家佛手”, 树势中等,生长与管理一致,株行距为2 m×3 m。在1998年,从7月4日开始,每隔15 d左右取叶样测定类黄酮、内源激素,摸索测定方法和了解其变化的大致规律1)。在1999年,从5月4日开始,每隔15 d左右于上午10:00,在36株银杏树上,分东、南、西、北4个方位,在生长势相同的同类枝条叶丛枝上随机采摘正常叶片100张,分3组,取样到11月29日止。采样后迅速将叶片洗净、晾干,经105 ℃杀酶20 min,65 ℃烘干至恒重、粉碎,干燥器贮存待测。用于测定内源激素含量的叶片样品,先液氮速冻,后放在-20 ℃冰箱中保存待测(程水源等, 2002)。
1) 程水源.影响银杏叶黄酮形成的主要因子及调控技术的研究.山东农业大学博士学位论文,2001
1.2 测定方法与数据图表处理类黄酮含量测定采用程水源等(2001)方法。乙烯测定1):从银杏叶丛枝上取叶片9张,称重,3次重复,密封于玻璃瓶内,4 h后用特制注射器抽取气体于岛津GC-14B型气相色谱仪测定ETH释放量,检测器FID,25 m×0.2 mm OV101柱,柱温105 ℃,进样口温度120 ℃,N2流速85 mL· min-1,H2压力0.6 kg· cm-2, 空气压力0.4 kg·cm-2, 外标法峰面积积分定量。采用酶联免疫法(ELISA)测定IAA、GA、ABA、异戊烯基腺嘌呤类(isopentengladenosines, iPAs)、玉米素类(zeatin ribosides, Zrs)含量,试剂盒由南京农大植物激素研究室提供。
试验处理及数据测定均重复3次,平均值表示结果,数据统计与制图采用软件DPS、Microsoft Excel。
2 结果与分析 2.1 银杏叶丛枝叶中类黄酮含量的动态变化银杏叶中类黄酮含量全年有2次高峰,第1次在7月4日(0.808 2%),第2次在11月1日(1.062 5%),第2次峰比第1次峰值提高了31.46%(图 1)。

|
图 1 银杏叶丛枝叶中类黄酮含量的动态变化 Fig. 1 The changes of flavonoids contents of leaves in G.biloba phyllome branch |
从表 1可知:IAA含量前期较高,后期尤其是行进落叶期较低,3次明显的含量高峰分别出现在5月18日、8月13日、9月29日,第1次峰值最高(198.74 nmol·g-1 FW),最低值在11月1日,仅为11.14 nmol·g-1 FW,最高值与最低值相差18倍。
|
|
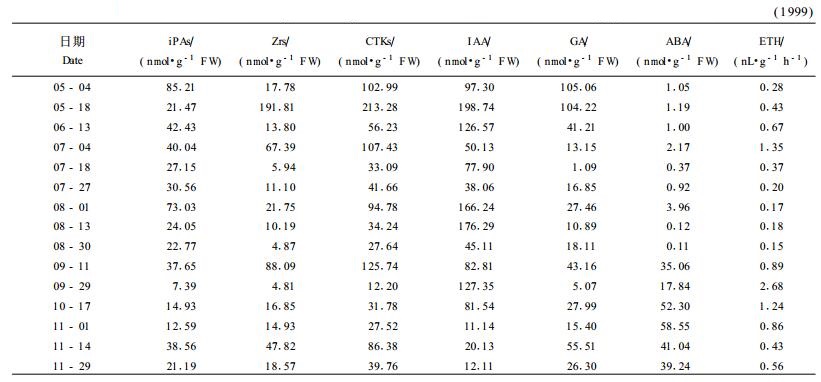
CTKs变化趋势与IAA基本一致,第1次峰与IAA同步,其他几次峰均提前半月左右。CTKs含量最高值与最低值相差也是18倍。GA含量变化与CTKs一致,但在5月上中旬出现含量高峰平台,幼叶是GA合成的主要场所,说明叶片GA此时期处于合成旺盛期。此后,GA持续下降直到7月中旬。最低含量(1.09 nmol·g-1 FW)与最高含量(105.06 nmol·g-1 FW)相差97倍,CTKs和GA含量变化曲线很相似,相关系数r=0.762 7**,达极显著水平。3种激素相比,IAA和CTKs含量绝对值较高,变幅较小,GA含量绝对值较低,起伏最大。Zrs和iPAs含量值分别处在4.81~191.81 nmol·g-1 FW、7.39~85.21 nmol·g-1 FW,变动幅度分别是40倍和11.5倍,说明在银杏叶片中Zrs的代谢强度比iPAs大。从变化趋势来看,Zrs与CTKs总量基本一致,二者相关系数高达0.916 3**。
综上所述,3种生长类激素在5月中旬之前含量最高,6、7月份是含量低谷期,8、9月份又有不同程度的含量高峰,10月份后含量均较低。其中,GA和IAA分别在早期和中期出现含量高峰平台,CTKs波动次数最多,GA含量变化幅度最大。
2.2.2 抑制类激素从表 1可知:ABA含量生长后期较高,生长前期和中期含量较低或甚微。具体表现为:在5月初ABA含量仅为1.05 nmol·g-1FW,在8月底之前含量绝对值一直维持在较低的水平,此后ABA含量迅速上升,于9月中旬和11月初分别达到全年次大值(35.06 nmol·g-1 FW)和最大值(58.55 nmol·g-1 FW),最高值与最低值相差532倍。其中,在7月初和11月初的ABA含量峰分别与叶中类黄酮含量峰相平行。
乙烯生成量峰分别在7月4日和9月29日,后一次峰值较大,最大值与最小值相差18倍。与ABA相比,在7月初,二者同时达到第1次高峰,在8月底二者同时进入含量高峰期,ABA先于乙烯达到较高值,但在其含量波动时,乙烯达到全年生成高峰,后ABA含量转为持续上升,最高峰滞后于乙烯1个月。后期高浓度的ABA、乙烯与叶片迅速成熟衰老有关(程水源等,2000;Max, 1990;Satoru et al., 1991;Sylvia et al., 1998)。与叶中类黄酮含量峰相比,第1次峰平行,第2次峰超前。
总之,生长素类激素在前期相对较高,后期较低,抑制类激素在前期和中期相对较低,后期较高。变化幅度ABA最大(532倍),GA其次(97倍),IAA、CTKs、ETH(18倍)最小,其ABA、ETH含量峰与叶中类黄酮含量峰有较好的一致性。
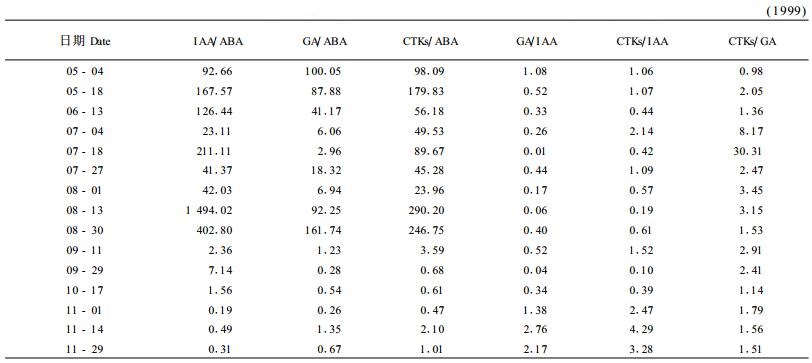
2.2.3 激素平衡数据转换结果如表 2。IAA/ABA、GA/ABA、CTKs/ABA表现出较大的绝对值水平,前期(6月中旬前)和中期(8月中、下旬)相对较大,后期较小,其比值大部分时候小于1,这说明后期ABA含量水平逐渐接近甚至超过了IAA、GA、CTKs。由于8月中下旬是IAA和CTKs较旺盛合成期,同时也是ABA含量的最低谷,因此IAA/ABA、CTKs/ABA比值最大。与图 1对比可知,后期IAA/ABA低谷正好与叶中类黄酮含量高峰同步,而GA/ABA、CTKs/ABA低谷则超前。
|
|
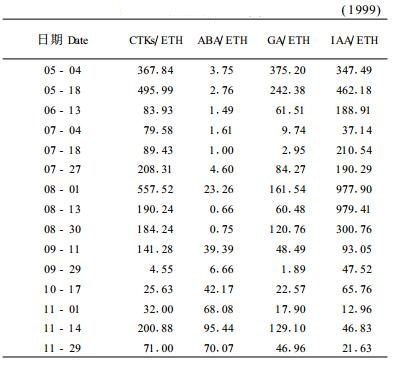
从表 3可知:CTKs/ETH、IAA/ETH、GA/ETH值早期较大,7月上、中旬下降到最低值,再后的变化不尽相同,但都于9月底又降到最低值或较低值。综上所述,生长类激素与ABA、ETH比值在7月初、9月底至11月初处于低谷,此时正是叶中类黄酮合成高峰期,其IAA/ABA、GA/ABA、IAA/ETH、GA/ETH值的低谷期与叶中类黄酮的高峰期在时间上同步。
|
|
银杏种子及花芽内源激素的研究己有报道,但叶片中相关研究尚未涉及1)(曹帮华等,2001)。通过对银杏叶中5大类6种激素分析可知,内源激素和叶中类黄酮之间关系密切。早期较高的乙烯生成峰促进叶中类黄酮合成的第1次高峰,此时ABA也伴随一次含量高峰,但绝对量较低。后期乙烯合成峰早于ABA启动,但随后ABA处于合成高峰,并维持在较高水平,相伴随出现的是叶中类黄酮全年合成最大峰。前人报道认为,ETH、ABA促进类黄酮(包括花青苷)合成(程水源等,2000;Goszybska et al., 1985;Max,1990;Satoru et al., 1991;Sylvia et al., 1998),并认为它们与调节酚类物质代谢有关(Apte,1982;Max,1990),本试验结果与之吻合。但本研究还说明ETH、ABA的作用大小有阶段性和主次之分,前期主要是由于乙烯的作用,ABA作用很小或仅起辅助作用,后期由于乙烯的启动,叶片走向成熟衰老,ABA含量激增,叶中类黄酮也大量合成,这是乙烯和ABA共同作用的结果,或乙烯诱导成熟,成熟衰老导致ABA剧增,ABA增加导致叶中类黄酮大量合成,即ABA是后期促进叶中类黄酮大量积累的直接原因。
1) 程水源.影响银杏叶黄酮形成的主要因子及调控技术的研究.山东农业大学博士学位论文,2001
GA含量高不利于类黄酮合成(Max, 1990),本试验结果与之大致类似。但从GA绝对量变化来看,也有不一致的地方,这可能与内源激素共同作用而不是由某一种激素单独起作用有关(曹帮华等,2001;王永章等,2000;Merlo et al., 1991)。
在早期和后期IAA含量由较高值陡降时,有利于叶中类黄酮合成,但在7月中旬至9月底之间有相反趋势:IAA从77.90 nmol·g-1FW陡降到38.06 nmol·g-1 FW,叶中类黄酮从0.712 4%下降到0.639 6%,IAA从176.29 nmol·g-1FW陡降到45.11 nmol·g-1 FW,叶中类黄酮从0.778 4%缓降到0.727 1%。本试验结果说明IAA与类黄酮的关系因时间不同而异。前期和后期IAA含量高,有利于叶片调集营养、进行光合作用和光合产物积累(郭维明等,1999;Max,1990),当IAA含量下降时,ETH含量增加,促进叶片成熟,次生代谢加强(王永章等,2000;Max,1990),叶中类黄酮迅速合成。中期则不然,或者由于这个阶段其它激素起更大作用,或者由于次生代谢向其它产物转化加强而影响了叶中类黄酮的合成。
在前期,CTKs的效果似乎与IAA作用相反,CTKs含量从低到高,叶中类黄酮合成积累加快,中后期则不然。有报道认为,CTKs促进类黄酮生成,并且这种作用是通过促进乙烯释放来完成的(Max,1990)。在叶片生长发育早期,本试验结果与之一致,但在中后期则不然,甚至相反。近年来,对CTKs中iPAs、Zrs的分析研究报道较多,它们对植物体生长发育的影响也不尽一致(曹帮华等,2001;Merlo et al., 1991)。本试验研究结果没有发现iPAs和Zrs与类黄酮之间的规律性关系。
总之,从内源激素含量变化的角度来看,ABA、ETH含量峰与叶中类黄酮含量高峰有较高的一致性,这说明2种激素在调控银杏叶中类黄酮合成中的重要性。
内源激素与叶中类黄酮之间的关系看来通过激素平衡来讨论更加合理。较小或最低的IAA/ABA、GA/ABA、CTKs/ABA有利于类黄酮的合成,即说明ABA相对含量较高比其绝对含量较高更有利于类黄酮合成,这对上述讨论中有些不太一致和矛盾的地方是一个很好的佐证。相对较高的CTKs/IAA有利于细胞的分化,而有文献证实,细胞分化期类黄酮合成旺盛(Lobstein,1991;Max,1990)。同样,CTKs/ETH、GA/ETH、IAA/ETH低,有利于类黄酮的合成,反过来说,相对较高的乙烯生成量促进类黄酮合成。ABA和乙烯同属促进类黄酮合成的激素(Max,1990),它们之间的比值与叶中类黄酮之间的关系不明显是可以理解的。仔细比较激素平衡与叶中类黄酮合成高峰之间的关系时发现:IAA/ABA、IAA/ETH、GA/ABA、GA/ETH值最小或接近最小时,叶中类黄酮含量最高。在离体培养时,有报道认为GA/ABA或GA/ETH影响类黄酮合成快慢,而不是某单一激素的作用(王永章等,2000)。本试验除证实激素平衡调控叶中类黄酮合成外,还找到了各种激素平衡对叶中类黄酮合成的具体影响:较高的CTKs/IAA,较低的CTKs/ABA、IAA/ABA、GA/ABA、CTKs/ETH、IAA/ETH、GA/ETH有利于叶中类黄酮形成,其中,以IAA/ABA、IAA/ETH、GA/ABA、GA/ETH的调控最为重要。在田间施加生长调节剂处理提高银杏叶中类黄酮的试验结果,进一步证实了上述结果的可靠性1)(王燕等,2002)。
1) 程水源.影响银杏叶黄酮形成的主要因子及调控技术的研究.山东农业大学博士学位论文,2001
内源激素促进类黄酮合成大多数是通过提高酶活性或促进成熟来完成的(郭维明等,1999;程水源等,2000;Max, 1990)。在银杏叶中类黄酮合成中,查尔酮合成酶(chalcone synthase, CHS)是最关键的酶,其次是苯丙氨酸氨解酶(L-phenylalanine ammonia-lyase, PAL)1)。通过对比激素含量及激素平衡变化可以发现:CHS酶受高含量IAA的刺激,较低的GA含量减少了对CHS酶和PAL酶的束缚,较高的乙烯加强了对CHS酶和PAL酶的诱导,相对较高的ABA含量拮抗了不利于类黄酮合成的激素如GA的影响1)(Carolyn et al., 1996;Max,1990;Satoru et al., 1991)等,这些因素的共同影响带来了类黄酮合成高峰。值得说明的是后期高ABA含量对酶活性的影响可能更大。因此,可以说内源激素含量及其平衡共同作用来调节关键酶的活性,影响叶片生长发育进程,调控叶中类黄酮合成。
曹帮华, 蔡青菊. 2001. 银杏种子生理研究进展. 山东农业科学, (1): 40-42. DOI:10.3969/j.issn.1001-4942.2001.01.024 |
程水源, 顾曼如, 束怀瑞. 2000. 银杏叶黄酮研究进展. 林业科学, 36(6): 110-115. DOI:10.3321/j.issn:1001-7488.2000.06.019 |
程水源, 顾曼如, 束怀瑞. 2001. 银杏叶黄酮含量变化与分布规律的研究. 园艺学报, (4): 353-355. DOI:10.3321/j.issn:0513-353X.2001.04.017 |
程水源, 王燕, 李俊凯, 等. 2002. 银杏叶中类黄酮类化合物合成代谢规律的研究. 林业科学, 38(5): 60-63. DOI:10.3321/j.issn:1001-7488.2002.05.010 |
郭维明, 盛爱武. 1999. 梅花采后衰老的内源激素的调节. 北京林业大学学报, 21(2): 42-47. DOI:10.3321/j.issn:1000-1522.1999.02.009 |
王燕, 程水源, 费永俊, 等. 2002. 提高银杏叶黄酮含量的调控措施. 湖北农业科学, (5): 103-105. DOI:10.3969/j.issn.0439-8114.2002.05.045 |
王永章, 张大鹏. 2000. 乙烯对成熟期新红星苹果果实碳水化合物代谢的调控. 园艺学报, 27(6): 391-395. DOI:10.3321/j.issn:0513-353X.2000.06.001 |
Apte P V. 1982. Inhibitory action of phenolic compounds on abscisic acid-induced abscission. J Expt Bot, 33: 826-830. DOI:10.1093/jxb/33.4.826 |
Carolyn E L, Jane E L, John R L. 1996. Developmental changes in enzymes of flavonoid biosynthesis in the skins of red and green apple cultivators. J Sci Food Agric, 71: 313-320. DOI:10.1002/(SICI)1097-0010(199607)71:3<313::AID-JSFA586>3.0.CO;2-N |
Goszybska D M, Rudnicki R M, Reid M S. 1985. The role of plant hormones in the postharvest life of cut flower. Acta Horticulturae, 167: 79-93. |
Lobstein A. 1991. Seasonal variations of the flavonoid content from Ginkgo biloba leaves. Planta Med, (57): 430-433. |
Max C S. 1990. External control of anthocyanin formation in apple. Scientia Horticulturae, 42: 181-218. DOI:10.1016/0304-4238(90)90082-P |
Merlo L, Passera C. 1991. The changes in carbohydrate and enzyme levels during development of leaves of Prunspersica. Plant Physiol, 73: 621-626. |
Satoru K, Jamnong U, Hiroshi G. 1991. Comparison of 1- aminocyclopane-1-carboxylic acid, abscisic acid and anthocyanin content of some apple cultivators during fruit growth and maturation. J Japan Soc Hort Sci, 60(3): 505-511. DOI:10.2503/jjshs.60.505 |
Sylvia M B, Richard C U. 1998. PAL and ethylene content during maturation of red and "Golden delicious" apples. Photochemistry, 27(4): 1001-1003. |