 2004, Vol. 40
2004, Vol. 40文章信息
- 任艳林, 张华荣, 李俊清.
- Ren Yanlin, Zhang Huarong, Li Junqing.
- 海南岛几种优势树种构筑型的研究
- Research on Architecture of Dominant Trees in Hainan
- 林业科学, 2004, 40(6): 39-44.
- Scientia Silvae Sinicae, 2004, 40(6): 39-44.
-
文章历史
- 收稿日期:2002-09-09
-
作者相关文章


由种子萌发生长成的幼树是一个分枝系统,幼树上所有的枝条都按照这个分枝体系进行分化,最后的生长发育结果便形成了树木基本的构筑模型结构(Halle et al., 1978)。
构筑模式在种和属的水平上规律十分明显,树木构筑模式与其属之间的关系已经很清楚。同一物种所有个体的发芽模式都是一样的(Keller, 1992),双子叶植物86%的属(Vogel, 1979)都有相同的特征。在树木发芽阶段,由幼苗所形成的主枝的形态往往不同于后来在树木主轴上观察到的特征(Kozlowski, 1971)。我国海南岛地处热带,树木每年的生长节律不明显,研究该地区树木的生长发育规律,从而根据生长特点采取合理的林业经营措施,就需要对这些树木的基本构筑模式进行精确的描述。
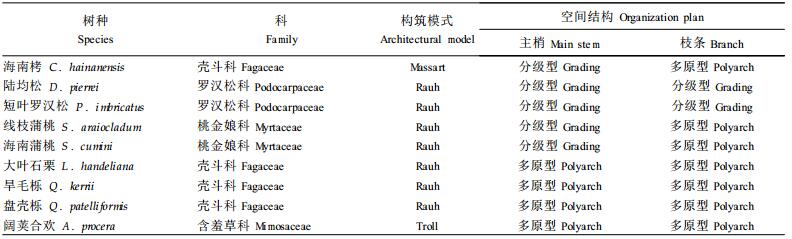
1 材料与方法 1.1 植物材料海南岛由于其地理隔绝的特点而为研究植物变异提供了很好的材料,本文选择了当地9种在林业上较重要的树种:海南栲(Castanopsis hainanensis)、线枝蒲桃(Syzygium araiocladum)、海南蒲桃(S. cumini)、陆均松(Dacrydium pierrei)、短叶罗汉松(Podocarpus imbricatus)、大叶石栗(Lithocarpus handeliana)、旱毛栎(Quercus kerrii)、盘壳栎(Q. patelliformis)、阔荚合欢(Albizzia procera)(Fu, 1992),见表 1。
|
|
树木茎轴是由一个芽顶端的分生组织产生的,整个树冠则可看作由一个或多个组织水平与多个分枝轴组成的体系。在这个体系中,根据分枝等级系列来区别枝条,任何一个等级系列的形成都与枝条的形状和生活力有关。在形态和构筑型特征分析时主要考虑了茎轴的生长节律、枝条形状及分枝模式。
树木的生长有一定的节律性,在连续的2个生长停滞期之间,茎轴生长1次,由此在2个生长期间形成生长单元(UG)。幼苗构筑型完全依赖第1个生长单元的形态特征,用4条简单的标准来描述该生长单元:胚轴长度,胚轴长则子叶位于土壤地表之上,胚轴短则子叶位于土壤之中;子叶特征,子叶为叶状或肉质;第一片叶的形态,鳞片状或叶状;叶序。
2 结果在所研究的9个树种中,树冠都由许多分枝茎组成,主干明显。茎枝节律性生长,并由一系列生长单元构成,侧生花序,节律性同龄分枝(侧枝与主枝同年生)。不同树种其树冠或为直生茎与斜生茎混合组成,或完全由同一种直生茎(斜生茎)组成(图 1)。

|
图 1 海南优势树种构筑型(主梢与基枝) Fig. 1 The architectural model of dominant trees in Hainan (main stem and base branch) |
海南栲树冠混生直生枝和斜生枝,主梢直生、单轴分枝,树枝斜生,单轴分枝或合轴分枝替代型,其生长模式是Massart型。在其它树种中,树冠全为直生茎或斜生茎:线枝蒲桃、海南蒲桃、陆均松、短叶罗汉松、大叶石栗、旱毛栎和盘壳栎均为直生茎,生长型符合Rauh型;阔荚合欢是斜生茎,生长型符合Troll型。
2.1 Massart型:海南栲树高约30 m,主干直生,枝条斜生(图 1-A)。树的主干直立式,辐射状对称,叶序螺旋状排列(图 2)。主干单轴分枝直生,决定整个树冠的构筑型,直生枝为分级型,主干上的基枝是斜生趋向水平的(图 2)。我们描述叶序的方式并不精确,只是肉眼观察到叶柄是扭曲的,因此有必要通过芽的解剖学研究来确定斜生枝及趋于斜生的直生枝。位于同一水平面上的主轴和侧轴会发生替代,主茎合轴分枝、替代,不利于侧枝生长。分级在不同茎之间很明显,基枝为多原型,在树冠中,上层枝条是斜生的,其特征是主梢与基生枝的中间类型。当主茎发生退化时,斜生或水平的枝条会替代它成为主干。树枝可能发生改变,直生枝条可变为斜生枝,直生枝、分级直立枝、斜生多原型的基生枝按茎序发生改变。所有的枝条分枝是异龄生长,有不同的生长单元,节律性生长,顶生(图 2)。

|
图 2 海南栲基枝 Fig. 2 The base branch of C. hainanensis |
树轴直生(图 1-B、C、D)。辐射状对称,叶序螺旋状排列,分枝异龄生长,生长单元节律性生长,顶生。
陆均松与短叶罗汉松高约30 m,除了在营养生长上的差异,主梢与树冠基部基本相似,分枝形态相同。在树冠顶部,生长单元较长,分枝较多,主梢和枝条按一定等级系列空间分布(图 3-A、B;图 4-A、B)。树枝常为单轴分枝直生(图 1-B)。陆均松的幼茎柔软狭长(图 3-A、B)。在主茎和侧枝上的分生组织或在2个生长单元间的组织明显变小变密。短叶罗汉松1年内的最后1个生长单元长出的针叶较大,最先发生的生长单元则布满了鳞片(图 4-A)。树木的主梢和幼苗在1年中最后1个生长单元分两行排列,叶柄扭曲后,向水平方向发展,逐渐发展成斜生枝。分生组织或2个生长单元之间的组织密被鳞状叶。

|
图 3 陆均松 Fig. 3 D. pierrei A.主枝The fleche; B.幼苗Juvenile individual. |

|
图 4 短叶罗汉松 Fig. 4 P. imbricatus A.幼苗Juvenile individual; B.基枝Base branch. |
线枝蒲桃和海南蒲桃树高10~25 m,在它们的主梢上,主干单轴分枝,直生(图 5-A)。与侧生枝相比,主干明显,生长旺盛,空间结构为分级型。基部的枝条和茎多为合轴分枝,这是在顶芽败育后由侧芽发育产生茎轴的形态特征(图 5-B)。侧枝与主干区分不明显,进而出现多原型空间排列。由分级型主梢形成的树干,主干单轴分枝,侧枝多原型(图 1-C)。所有枝条的叶序均为螺旋状排列,叶对生或交互对生,但有时三个一组排列在一个垂直面上(图 5-A、B)。花序及果序侧生于较老的生长单元上,叶腋脱落(图 5-C)。主干异龄生长,侧枝同龄生长。果卵形,两两对生(图 6-A、B)。

|
图 5 线枝蒲桃 Fig. 5 S. ariaocladum A.主枝The fleche; B.基枝The base branch; C.花序The inflorescence. |

|
图 6 盘壳栎 Fig. 6 Q. Patelliformis A.主枝The fleche; B.基枝The base branch; C.幼苗The juvenile individuals. |
大叶石栗、旱毛栎和盘壳栎主梢及树冠相似,形态相同,生活力有差别。生长单元长,分枝密,树冠顶端芽败育现象少于侧枝。叶序螺旋状排列,茎枝常短生且合轴分枝(图 1-D)。由几个生长单元发育而成的幼苗,具有单轴分枝的主干或分级型合轴分枝的空间结构(图 6-C)。老龄树的多原型组织出现要晚得多。
2.3 Troll型:阔荚合欢树冠所有的茎轴为斜生(图 1-E)。主茎在叶落后成直立。叶序对生排成2列(图 7-A, B)。形状简单,分枝节律性,顶生,异龄生长。

|
图 7 阔荚合欢 Fig. 7 A. procera A.主枝The fleche; B.基枝The base branch; C.花序The inflorescence. |
在树冠上部,主枝都是由许多顶端分生组织发育而成(图 7-A)。树枝扭曲,在叶子背面成之字形,相对于斜生枝主茎生长衰弱。在同一水平面上,直生枝为多原型。在树冠下部,树枝功能不分化,为多原型(图 7-B)。枝条合轴分枝,替代型生长,扭曲或成之字形,花产生在枝条顶端。所有的花或果都是侧生(图 7-C)。花轴全为同龄生长,花序为向心形、长条形、圆锥形。
3 讨论 3.1 构筑模式与属海南树木与已研究过的植物属比较,其构筑型与属之间的关系更加确定。海南栲为Massart型,与欧洲和日本的栗属和栲属相同(Bourgeois, 1993; Soumoy et al., 1996);陆均松属于Rauh型,与新西兰同属的其他树种相同(Philipson, 1988);短叶罗汉松属于Rauh型,与柳叶罗汉松(Podocarpus salicifolius)相同(Halle et al, 1978);大叶石栗属于Rauh型,与Lithocarpus teysmanii相同(Halle et al, 1978);旱毛栎、盘壳栎属于Rauh型,与北美、欧洲、日本对栎属其它种的研究相同(Halle et al, 1978; Soumoy et al., 1996);阔荚合欢属于Troll型,与对合欢(A. julibrissin)、大叶合欢(A. lebbek)的研究相同(Halle et al, 1978)。
在所研究的植物中,有7种是Rauh型,1种是Massart型,1种是Troll型(表 1)。在主梢和基生枝中,有5种为分级型直生枝,4种为多原型直生枝;有2种枝条为分级型,其余7种为多原型。在Rauh型中有生长的分化,分为主梢与基枝均为分级型(陆均松和短叶罗汉松)、主梢分级型枝条多原型(线枝蒲桃和海南蒲桃)以及主梢与枝条均为多原型(大叶石栗、旱毛栎和盘壳栎)。
3.2 发芽模式与属海南栲发芽模式是子叶留土,与栲属的C. foxworthyi、C. inermis、C. javanica、C. berangau(NG, 1980, 1991), 尖叶槠(C. cuspidate)(Soumoy et al., 1996), 欧洲栗(Castanea sativa)相同(Csapody, 1968; Muller, 1978);陆均松发芽模式是子叶出土,与柏木陆均松(Dacrydium cupressinum)相同(Philipson, 1988);短叶罗汉松发芽模式是子叶出土,与百日青(Podocarpus neriifolius)(NG, 1978)、新西兰罗汉松(P. totara) (Philipson, 1988)、P. coriaceus(Duke, 1965)相同;线枝蒲桃和海南蒲桃发芽模式是子叶留土,与蒲桃属(Duke, 1969;Miquel, 1987)、蒲桃(S. jambos)(Rousteau, 1983)相同;大叶石栗发芽模式是子叶留土,与粗穗柯(L. elegans)、L. encleisacarpus、L. ewyckii、L. mempening相同(NG, 1978;1991);旱毛栎和盘壳栎发芽模式是子叶留土,与Q. oidocarpa、Q. mempening(NG, 1991)、苦栎(Q. cerris)、黎巴嫩栎(Q. libani)、无梗栎(Q. petreae)、柔毛栎(Q. pubescens)、欧洲白栎(Q. robur)、北美红栎(Q. rubra)(Csapody, 1968;Muller, 1978)、日本常绿栎(Q. acuta)、麻栎(Q. acutissima)、槲树(Q. dentata)(Soumoy et al., 1996)相同;阔荚合欢发芽模式是子叶出土,与大叶合欢相同(Duke, 1965; Schopmeyer, 1974; Miquel, 1987)。
3.3 由直生枝向斜生枝的转变由直生枝向斜生枝的转变在海南研究的9种树中只有2种,即海南栲和阔荚合欢,其生长型分别为Massart型和Troll型。发芽过程中,主茎的第1个生长单元直生。1个生长周期后,第2个生长单元开始斜生,如合欢属侧枝则从第1个生长单元开始时就斜生。这种茎轴的转换是在第1个生长停滞期开始的。如同温带树种一样,留下了冬芽痕迹,这种转变会年复一年地进行,如Massart型中的欧洲栗和米槠(Castanopsis cuspitata),Troll型中的欧洲水青冈(Fagus sylvatica)、日本山毛榉(F. crenata)、米心水青冈(F. engleriana)、美国水青冈(F. grandifolia)、日本水青冈(F. japonica)、水青冈(F. longipetiolata)、亮叶水青冈(F. lucida)、东方水青冈(F. orientalis)、巴山水青冈(F. pashanica)、假水青冈属的Nothofagus alessandrii、N. alpina、N. betuloides、N. glauca、N. nitida、N. pumilio、南极假水青冈(N. antartica)、南方假水青冈(N. dombeyi)、歪叶假水青冈(N. oblique)(Soumoy et al., 1996; Thiebaut et al., 1997)。在所有树种中,第1次停止生长不仅是1种生长节律,而且是1个茎轴的转变过程。
3.4 构筑型与林业本文所研究树种构筑型的共同特征是:主干区别于其他枝条,生长具节律性,花序侧生,分枝节律性且顶生。当然,并不是每一种树在林业上的构成方式都一样,它们有不同的特征,例如茎直生或斜生,主茎及空间结构为分级型或多原型,这些区别将造成林业管理措施的不同。
培养一株树干通直的树木,在其树冠是分级型的情况下,主茎突出,个体独特性强,林业经营就简单;而树冠为多原型时,主茎不明显,个体独特性不强,林业经营就复杂。同样,当树木的主茎是直生型而且相互依赖时,就相对容易形成通直高大的树干,而主茎是斜生时,斜生枝需要一个直立的过程,培养通直高大的树干就十分复杂。至于构筑型与林业经营的确切关系,还需要进一步研究。
Bourgeois C. 1993. Le Châtignier, un arbre, un bois. Institut pour le d veloppement forestier, Paris, France. |
Csapody V. 1968. Keimlings Bestimmungsbuch der Dikotyledonen. Akademia Kiado, Budapest. |
Duke J A. 1965. Keys for the identification of seedlings of some prominent woody species in 8 forest types in PuertoRico Ann. Missouri Bot Gard, 52: 314-350. DOI:10.2307/2394796 |
Duke J A. 1969. On tropical tree seedlings. I seeds, seedlings, systems and systematics. Ann. Missouri Bot Gard, 56: 125-161. DOI:10.2307/2394836 |
Fu S. 1992. List of trees in the arboretum of Jianfengling. Publication of the forest station Jianfenglin, Hainan: 76. |
Halle F, Oldeman R A A, Tomlinson P B. Tropical trees and forests-an architectural anylysis. Springer-Verlag, Berlin, Heidelberg, 1978
|
Keller R. Les familles des Dictyledones ligneuses tropicales et leur identification a l'aide d'une clef fondee sur les seuls caract res v g tatifs. These de doctorat.Universite de Montpellier 2, Montpellier, 1992
|
Kozlowski T T. Growth and development of trees. I Seed germination, ontogeny and shoot growth. Academic Press, New York, 1971
|
Miquel S. 1987. Morphologie fonctionnelle de plantules d'especes forestieres du Gabon. Bulletin Museum National d'Histoire Naturekke, Pairs, 4e serie, 9section b, Adansonia, 1: 101-102. |
Muller F M. Seedlings of the north-western european lowland. A flora of seedlings. Dr. W Junk-B V. Publishers, Boston, 1978
|
NGFSP. Strategies of establishment in Malayan forest trees. In Tomlinson P B & Zimmermann M H(eds), Tropical trees as living systems. Cambridge University Press, Cambridge, 1978: 129-162
|
Philipson W R. 1988. Seedling and shoot morphology of the New Zealand species of Nothofagus (Fagaceae). N Z J Bot, 26: 401-407. DOI:10.1080/0028825X.1988.10410643 |
Rousteau A. 1983. 100 plantules d'arbres guadeloupeens. Aspects morphologiques et ecologiques Universite Pares 6, dilome de docteur 3 e cycle: 100. |
Schopmeyer C S. 1974. Seeds of woody plants in the United States. US Dep Agric Handb: 450. |
Soumoy L, Thiebaut B & Serey I. 1996. D veloppement de quelques Fagaceae japonaises; germination, modèle architectural et syst matique. Can J of Bot, 74: 1248-1259. DOI:10.1139/b96-151 |
Thiebaut B, Serey I, Druelle J L, et al. 1997. Forme et architecture de quelques hêtres, chiliens (Nothofagus Blume) et chinois (Fagus L.). Can J of Bot, 75: 640-655. DOI:10.1139/b97-072 |
Vogel E F. Seedlings of dicotyledons. Centre of Agricultural Publishing and documentation, Wageningen, 1979
|