 2004, Vol. 40
2004, Vol. 40文章信息
- 闫淑君, 洪伟, 吴承祯.
- Yan Shujun, Hong Wei, Wu Chengzhen.
- 福建万木林中亚热带常绿阔叶林林隙更新研究
- Gap Phase Regeneration in Mid_Subtropical Evergreen Broad-Leaved Forest in Wanmulin, Fujian
- 林业科学, 2004, 40(6): 25-31.
- Scientia Silvae Sinicae, 2004, 40(6): 25-31.
-
文章历史
- 收稿日期:2003-11-06
-
作者相关文章


2. 福建农林大学林学院 福州 350002
2. Forestry College of Fujian Agriculture and Forestry University Fuzhou 350002
林隙(gap)可分为冠空隙和扩展林隙2类。冠空隙是指直接处于林冠层空隙下的土地面积或空间;扩展林隙是指由冠空隙周围树木的树干所围成的土地面积或空间,它包括了实际林隙和其边缘到周围树木树干基部所围成的面积或空间2部分(Watt,1925;1947;Runkle, 1981;1982)。树倒的发生和林隙的形成,使林隙内的环境发生了很大变化,随着这些环境条件的改变,林隙内的生物种类组成、群落结构与功能也与林内显著不同(Brokaw,1987;Lawton,1990;Tyrrell,1994;臧润国等,1999a;1999c;王周平等,2001;2003)。不同树种对林隙环境的占据和利用特性不同,从而使其更新在林隙内外表现出一定的差异(罗大庆等,2002;付士磊等,2003)。
我国分布着世界上独有的面积最大的亚热带常绿阔叶林,是世界上发生古老、类型丰富、结构复杂的极具代表性的植被类型,是植物群落和生物多样性保护的关键地区之一(Kloetzli,1981;李昌华,1997;吴征镒,1980)。了解常绿阔叶林林隙更新及其特点、不同树种更新行为差异,对常绿阔叶林多样性保护和维持有重要的现实意义。在对中亚热带常绿阔叶林林隙边缘木高径分布特征、林隙面积的Weibull分布以及林隙内主要树种高度生态位研究的基础上(闫淑君等,2002;2003),现针对福建万木林中亚热带常绿阔叶林扩展林隙的更新动态进行初步研究。
1 研究地点与研究方法 1.1 研究地点的自然概况调查地点设在福建省北部万木林自然保护区,即118°08′22″—118°09′23″ E,27°02′28″—27°03′32″ N。全区南北长1.82 km,东西宽1.04 km,土地总面积1.89 km2。属中亚热带季风型气候,水、光、热资源丰富,年平均气温18.7 ℃,年平均降雨量为1 663.8mm,土壤为红壤。万木林原为元末富绅杨达卿营造的一片杉木(Cunninghamia lanceolata)人工林,历经600多年的封禁保护,经过了几个世纪的次生演替重新恢复为地带性植被——中亚热带常绿阔叶林,而成为具有原始森林景观的自然群落。它代表着亚热带温暖湿润地区的常绿双子叶植物阔叶树林,主要由壳斗科(Fagaceae)、樟科(Lauraceae)、山矾科(Symplocaceae)、山茶科(Theaceae)、木兰科(Magnoliaceae)、金缕梅科(Hamamelidaceae)和杜英科(Elaeocarpaceae)的常绿阔叶树种组成。
1.2 研究方法采用样线调查法(Runkle,1982)。从样线起点开始调查每个林隙,测其扩展林隙的长轴和短轴,以椭圆形测算林隙面积。根据林隙面积把林隙分为7个等级,即:<100 m2, 100~200 m2, 200~300 m2, 300~400 m2, 400~500 m2, 500~600 m2, ≥600 m2。每个等级的林隙选2个,共调查林隙14个,同时,分别在每个林隙中均匀地设置5个4 m×4 m的小样方,并在离林隙边缘约10 m远处随机设置4个5 m×5 m的非林隙区样方。分别调查林隙内及非林隙区的乔木、灌木的种类、个体数量、胸径及高度等。同一大小级2个林隙的平均物种个体数作为该等级的物种个体数。每个物种在单位林隙面积上的个体数称之为该物种的更新密度。
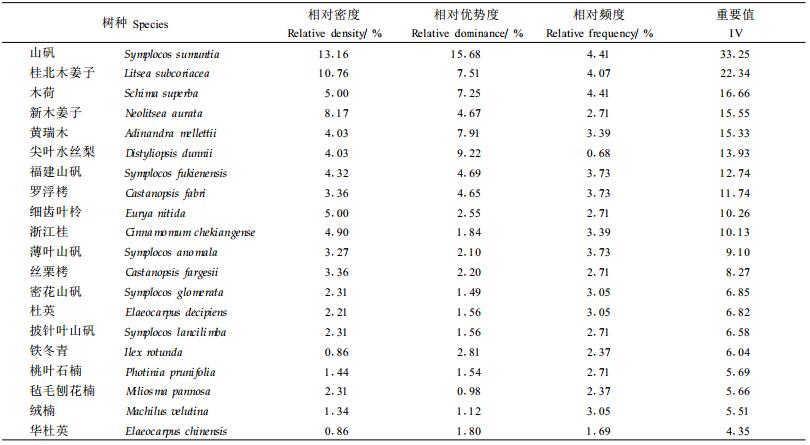
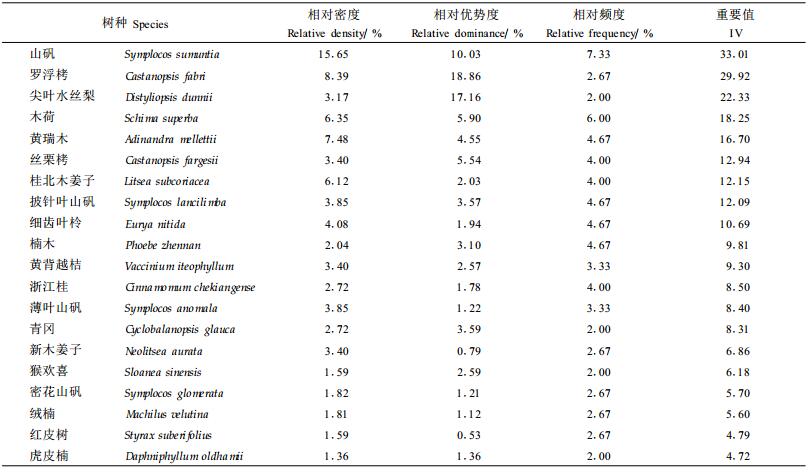
2 结果与分析 2.1 乔木树种在林隙内外的数量特征对比分析在林隙内共调查到乔木树种75种(1.5 m以上的),而在非林隙中有52种。由于中亚热带常绿阔叶树种繁多,仅列出了20个主要树种在林隙和非林隙中的数量特征,作为说明示例。不同树种在林隙和非林隙林分中出现的频度、密度和显著度不同,从而表现出其重要值的不同(表 1、2)。林隙内以山矾(Symplocos sumuntia)、桂北木姜子(Litsea subcoriacea)、木荷(Schima superba)最占优势,而在非林隙中则是山矾、罗浮栲(Castanopsis fabri)和尖叶水丝梨(Distyliopsis dunnii)最占优势。为了分析各树种在林隙和非林隙林分内优势度的变化规律,分别对林隙内和非林隙内的所有树种按其重要值大小进行了位序排列,对每个树种都给出了其在林隙内的的重要值位序(Og)和在非林隙林分中的重要值位序(Ong)(表 3)。无论在林隙内还是在非林隙林分中,山矾的重要值都位居第1,也就是说山矾是最大程度地利用了林隙环境并迅速成长,直到林冠层的树种。在从林隙到林冠转变过程中,山矾始终处在优势地位。山矾在非林隙中处主导地位,正说明山矾是群落建群种;山矾在林隙内占优势,说明它是典型的林隙更新方式。而黄瑞木(Adinandra mellettii)在林隙内和林隙外都位居第5位,细齿叶柃(Eurya nitida)都位居第9位,说明二者对林隙的反应不明显。其它的树种在林隙内外的重要值的次序都有所变动。
|
|
|
|
|
|

不同树种在林隙内和非林隙林分中的优势地位不同,同一树种在林隙内和非林隙中的优势地位也有所不同。为了便于分析和表达,规定:只在林隙内出现,而在非林隙林分内不出现或很少出现的树种,称其为林隙树种(gap species); 而只在非林隙中出现,在林隙内不出现或很少出现的树种,称其为非林隙树种(non-gap species),这类树种有不少是只有在密闭阴暗的林下才能更新的树种;在林隙内的重要值位序先于非林隙中位序(即Og-Ong<0)的树种称其为对林隙产生正更新反应的树种,将在林隙内的重要值位序后于非林隙中位序(即Og-Ong>0)的树种称为对林隙产生负更新反应的树种;将︱Og-Ong︱>10以上的树种,规定为林隙更新反应强烈的树种,将︱Og-Ong︱<5的树种,规定为对林隙更新反应不明显的树种,而将5≤︱Og-Ong︱≤10之间的树种,规定为对林隙更新反应中等的树种(臧润国等,1999a)。根据上述规定,则万木林中亚热带常绿阔叶林对林隙更新反应的树种组分别是:(1)林隙树种薄叶山矾、桃叶石楠、华杜英、观光木(Tsoongiodendron odorum)、华南桂(Cinnamomum austro-sinense)、香叶树(Lindera communis)、台湾冬青(Ilex rotunda)、拉氏栲(Castanopsis lamontii)、榕叶冬青(Ilex ficoidea)、黄檀(Dalbergia hupeana)、拟赤杨(Alniphyllum fortunei)、树参(Dendropanax dentiger)、福建冬青(Ilex fukinensis)、石栎(Lithocarpus glaber)、苦槠(Castanopsis sclerophylla)、枫香(Liquidambar formosana)、东南野桐(Mallotus lianus)、泡桐(Paulownia fortunei)、乌桕(Sapium sebiferum)、污毛粗叶木(Lasianthus hartii)、赤楠(Syzygium buxifolium)、乐东南拟单性木兰(Parakmeria lotungensis)、豆梨(Pyrus calleryana)、厚皮香(Ternstroemia gymnanthera)、闽楠(Phoebe bournei)、薯豆(Elaeocarpus japonicus)、罗浮柿(Diospyros morrisiana)、刨花楠(Machilus pauhoi)和山苍子(Litsea cubeba)。(2)非林隙树种弯蒴杜鹃(Rhododendron henryi)、大叶冬青(Ilex latifolia)、油柿(Diospyros oleifera)、山茶(Camellia japonica)和格药柃(Eurya muricata)。对表 3所列出的树种分析其对林隙更新的反应,(1)对林隙有强烈正更新反应的树种有:新木姜子、福建山矾、杜英和毡毛刨花楠;(2)对林隙有强烈负更新反应的树种有:楠木和黄背越桔;(3)对林隙有中等正更新反应的树种有:桂北木姜子、密花山矾和铁冬青;(4)对林隙有中等负更新反应的树种有:罗浮栲、丝栗栲和披针叶山矾;(5)对林隙更新反应不显著的树种有:山矾、木荷、黄瑞木、尖针水丝梨、细齿叶柃、浙江桂、薄叶山矾、绒楠和红皮树。
2.2 灌木对林隙的反应灌木树种对林隙也存在一定的反应,有的灌木在非林隙中没有调查到,如青灰叶下珠(Phullanthus glaucus)、盐肤木(Rhus chinensis)和血党(Ardisia brevicaulis)。每种灌木在林隙内的密度都非常明显地高于非林隙中,林隙内每种灌木的密度是非林隙中同种灌木密度的1.6~11.4倍左右,林隙内所有灌木的总密度为22 374株·hm-2,而非林隙中所有灌木的总密度为9 934株·hm-2(表 4)。林隙内大多数灌木的平均高度也小于非林隙中。这是由于林隙内的个体大多是林隙形成后形成的,年龄较小,所以大多高度还较小,而林下的灌木则大多是在林下生存了较多年的个体,其每个个体的平均就较高(表 4)。所以,林隙的形成,有利于灌木树种个体数量的增加、密度的增大、生长的加快。
|
|

林隙大小是林隙的重要特征之一,它影响着林隙内的环境因子及其组合状况以及资源对植物的有效性。通过生态因子的影响,林隙大小对各种植物的生长和繁殖产生不同的作用,进而影响到各种植物在林隙内的优势程度以及种间关系的变化。林隙的大小不同,反映林隙环境与周围非林隙环境的差别程度。所以,林隙研究者常以林隙大小作为林隙干扰的重要特征来研究树种的更新反应规律(Runkle, 1981;1982)。万木林中亚热带常绿阔叶林主要树种在不同大小林隙内的平均密度的变化情况,见图 1。

|
图 1 6个树种的平均更新密度随林隙大小的变化 Fig. 1 The regeneration response of 6 tree species to gap size |
各树种对林隙大小的反应虽然各不相同,但也表现出一定的共性:(1)各树种的更新密度总趋势都是随着林隙面积的增加呈下降;(2)100 m2左右的林隙中,山矾和桂北木姜子的平均更新密度趋于最大,而随着林隙面积的增大,表现出比较明显的下降趋势,但在林隙面积为500~600 m2时又有峰值出现;(3)木荷在林隙面积为300~400 m2时,其平均更新密度出现很明显的峰值;(4)新木姜子、黄瑞木和细齿叶柃三者都是在林隙面积为100~200 m2时,其平均更新密度出现明显的峰值,而细齿叶柃在300~400 m2又出现一峰值,黄瑞木在500~600 m2时出现一小峰值。100~200 m2和500~600 m2的林隙,是树种更新密度形成峰值的两个区域,尤其是100~200 m2,是树种平均更新密度发生变化的一个敏感区域,这与南亚热带常绿阔叶林有所不同的是,在南亚热带500 m2左右林隙是树种平均更新密度发生变化的一个敏感区域(臧润国等,1999c);除山矾外,其他树种在林隙面积<100 m2时,其更新密度较小,而在100~200 m2的林隙中更新密度达到最大,木荷则是林隙面积为300 m2左右时达到最大,在以后各大小级林隙中平均密度基本上呈逐渐下降的趋势。这种反应的形成原因可能是:在<100 m2的林隙内,更新环境有所改善,资源有效性比林下增大,所以其中的密度大于林下。但在<100 m2的林隙内的环境条件仍受周围大树的影响较大,尤其是在常绿阔叶林中,因为常绿阔叶林的树种多冠幅较大,林冠对光的遮挡以及周围大树根系对水分、养分的争夺等因素对林隙内更新树种仍有较大的作用,即以<100 m2的林隙内,大多数树种的更新密度不会太大。当林隙为100~200 m2时,林隙内的环境条件和资源有效性比林下有明显的改善,从而促进了树种的大量更新,同时较大的林隙面积内,草灌木也不太繁茂,它们对更新树种的竞争作用还较小,没有对更新形成较大的阻碍作用,所以,这么大的林隙内,大多数树种的更新密度最大。但当林隙面积大于200 m2以后,林隙内的环境空间和资源有效性比林下更加增大,特别是光照条件十分充足,而这样的条件却又大大地促进了草灌木的大量繁殖,这些草灌木很快就会占据林隙中的地面空间,覆盖度大增加,这样,对大多数乔木树种的更新就起到了抑制作用。所以随林隙面积的进一步增加,乔木树种的更新密度却大大减少了。而木荷最大更新密度出现在林隙面积为300~400 m2时,这与其生物学特性有关,因木荷是喜光树种,幼年较耐庇荫而喜上方光照,在草灌木的竞争压力下,因其特性使其在较大的林隙中仍有较好的更新。可见,树种在林隙内的平均更新密度,既受周围大树的影响,又受到林隙内竞争草本、特别是灌木的作用。总之,树种的更新对林隙大小的反应是各树种的综合特性对林隙内复杂的无机和有机环境因子复合体的系统反应,因此,在分析树种的更新对林隙大小的反应时,既要考虑到树种的耐荫性、繁殖力等生活史对策,又要考虑到不同大小林隙内的无机和有机环境因子复合体。
3 小结与讨论本研究林隙内的样方面积为80 m2,林隙外的样方面积为100 m2,主要原因是考虑到有小于100 m2的林隙必须研究,同时万木林中亚热带常绿阔叶林群落目前较为稳定(洪伟,1999),林隙外样方面积为80 m2与100 m2的多样性几乎没有变化。
重要值的大小所表明的是一个树种在群落中的优势地位,它在一定程度上表明了一个种相对于群落中其它种对生态资源的占据和利用能力的大小。在林隙中重要值越大,重要值位序越居前的树种,它们对林隙中生态资源的利用和竞争能力越强。与非林隙林分相比,林隙内的光照明显增大,林隙内气温、湿度、土壤温度等小气候因子的日变化和年变化规律就会发生相应的改变,土壤有机质分解速率和养分的有效性等也会发生不同程度的变化,同一物种由于林隙内外生态环境的不同,其在林隙内外的优势度也有所不同。随着林隙内生态因子的变化,不同生态特性的树种就会在林隙内有不同的更新和生长,从而使不同的树种在林隙内具有不同的优势度。万木林中亚热带常绿阔叶林中林隙内是以山矾、桂北木姜子和木荷为优势种,而在非林隙林分中是以山矾、罗浮栲和尖叶水丝梨为优势种。根据不同树种在林隙内外的重要值位序的差值大小,将万木林中亚热带常绿阔叶林树种对林隙干扰的反应分为5类:(1)对林隙有强烈正反应的树种有:新木姜子、福建山矾、杜英、毡毛刨花楠;(2)对林隙有强烈负反应的树种:楠木、黄背越桔;(3)对林隙中等正反应的树种有:桂北木姜子、密花山矾、铁冬青;(4)对林隙中等负反应的树种有:罗浮栲、丝栗栲、披针叶山矾;(5)对林隙反应不明显的树种有:山矾、木荷、黄瑞木、尖针水丝梨、细齿叶柃、浙江桂、薄叶山矾、绒楠和红皮。
万木林中亚热带常绿阔叶林中,大部分灌木在林隙内的更新密度均显著大于林隙外非林隙的,大多数灌木在林隙内的平均高度小于非林隙内的。林隙内的灌木比非林隙中多的原因主要是:由于林隙形成前,灌木上层的大树即林隙形成木,强烈地抑制着灌木的繁衍和生长,林隙形成木倒下或死亡后,抑制灌木的大树移去了,林隙内的可利用空间增大,光和其他可利用的资源也增加,在环境资源可利用程度增加的情况下,它们能快速地由无性繁殖或种子库中的种子产生新个体,新个体和原有的个体在有利环境资源下都能快速生长,所以,与非林隙相比,灌木的密度增加了,其总生物量也增加了。在林隙形成的初期灌木树种繁茂生长,得到了良好的发育,它们强烈地抑制乔木的更新和幼苗的生长,随着时间的推移,林隙的填充(侧生长与更新生长填充)程度也不断增加,灌木的密度和高度等就会逐渐接近最大极限,部分个体开始衰亡,林隙内的环境受灌木的其它树种的改造,再加上周围树木侧生长的作用,使林隙内的环境反而不利于灌木的生长和繁殖,却会对一些繁殖和生长快的中小乔木树种如桂北木姜子、新木姜子和黄瑞木等有利,乔木树种的繁茂进一步抑制了灌木的生长和繁殖。随着林隙的进一步填充和发育,灌木的密度就会因大部分个体的死亡和新个体的生长率和繁殖率降低。随着大部分灌木个体退出林隙,林隙环境有利于一些大乔木树种如木荷、罗浮栲和浙江桂等在林隙内更新。随着林隙的进一步演替,其内树木的高度和周围主林层接近时,林隙就逐渐消失,林隙斑块发育成熟林分,即林隙和周围林分没有多大的差别。这时,灌木的密度较林隙内的小,小乔木如桂北木姜子、新木姜子和黄瑞木等在林分中的重要性或优势度也较林隙中降低,而大乔木树种的重要性和优势度却逐渐增大,直到原来的林隙斑块发育到衰退阶段,受外力干扰重新形成新林隙后,灌木和小乔木的优势度才会又一次增大,从而进入下一轮森林循环过程。
目前对万木林中亚热带常绿阔叶林林隙生态环境随着林隙面积增加的变化规律还没有做过测定和分析,对不同树种的生态学特性也知之甚少,所以,在分析研究结果的机理时就显得论据不足。只有深入了解不同树种的生态学,特别是其生理生态学特性,并将树种物的特性同林隙生态环境以及群落中种间关系的变化规律紧密地结合起来进行综合分析,才能真正了解中亚热带常绿阔叶林众多树种共存和多样性维持的奥秘。本研究结果为在中亚热带常绿阔叶林中进一步有针对性的开展林隙更新机理的研究奠定了基础。
付士磊, 刘兴双, 宋晓东, 等. 2003. 长白山自然保护区阔叶红松林林隙更新状况. 辽宁林业科技, (6): 1-3. DOI:10.3969/j.issn.1001-1714.2003.06.005 |
洪伟. 1999. 闽江流域森林生态研究. 厦门: 厦门大学出版社, 105-121.
|
李昌华. 1997. 常绿阔叶林在东亚的分布. 自然资源, (2): 37-45. |
罗大庆, 郭泉水, 薛会英, 等. 2002. 西藏色季拉山冷杉原始林林隙更新研究. 林业科学研究, 15(5): 564-569. DOI:10.3321/j.issn:1001-1498.2002.05.010 |
罗大庆, 郭泉水, 薛会英, 等. 2002. 藏东南亚高山冷杉林林隙特征与干扰状况研究. 应用生态学报, 13(7): 777-780. DOI:10.3321/j.issn:1001-9332.2002.07.003 |
王周平, 李旭光, 石胜友, 等. 2001. 重庆缙云山针阔混交林林隙树木更替规律研究. 植物生态学报, 25(4): 399-404. DOI:10.3321/j.issn:1005-264X.2001.04.003 |
王周平, 李旭光, 石胜友, 等. 2003. 缙云山森林林隙与非林隙物种多样性比较研究. 应用生态学报, 14(1): 7-10. DOI:10.3321/j.issn:1001-9332.2003.01.002 |
吴征镒. 1980. 中国植被. 北京: 科学出版社, 1375.
|
闫淑君, 洪伟, 吴承祯, 等. 2002. 中亚热带常绿阔叶林林隙面积的Weibull分布模型研究. 江西农业大学学报, 24(6): 802-805. DOI:10.3969/j.issn.1000-2286.2002.06.016 |
闫淑君, 洪伟, 吴承祯, 等. 2002. 万木林中亚热带常绿阔叶林林隙主要树种的高度生态位. 应用与环境生物学报, 8(6): 578-582. DOI:10.3321/j.issn:1006-687X.2002.06.002 |
闫淑君, 洪伟, 吴承祯, 等. 2003. 中亚热带常绿阔叶林林隙边缘木高径分布特征. 福建林学院学报, 23(1): 5-8. DOI:10.3969/j.issn.1001-389X.2003.01.002 |
臧润国, 余世孝, 刘静艳, 等. 1999a. 海南霸王岭热带山地雨林林隙更新规律的研究. 生态学报, 19(2): 151-158. |
臧润国, 徐化成, 高文韬. 1999b. 红松阔叶林主要树种对林隙大小及其发育阶段更新反应规律的研究. 林业科学, 35(3): 2-9. |
臧润国, 刘静艳, 董大方. 1999c. 林隙动态与森林生物多样性. 北京: 中国林业出版社.
|
Brokaw N V L. 1987. Gap phase regeneration of three pioneer tree in a tropical forest. J Ecology, 75: 9-19. DOI:10.2307/2260533 |
Kloetzli F. 1988. On the global position of the evergreen broad-leaved forest in the subtropical and temperate zones. Veroff Geobot Inst ETH Stiftung Ruebel Zurich, 98: 169-196. |
Lawton R O. 1990. Canopy gaps and light penetration into a wind-exposed tropical lower mountain rainforest. Can J For Res, 20: 659-667. DOI:10.1139/x90-088 |
Runkle, J R. 1981. Gap regeneration in some old-growth forests of the eastern United States. Ecology, 62(4): 1041-1051. DOI:10.2307/1937003 |
Runkle, J R. 1982. Pattern of disturbance in some old-growth mesic forests of eastern North America. Ecology, 63: 533-546. |
Tyrrell LE, Crow T R. 1994. Structural characteristics of old-growth hemlock-hardwood forests in relation to age. Ecology, 75: 370-386. DOI:10.2307/1939541 |
Watt AS. Pattern and process in the plant community. J Ecology, 1947
|