 2004, Vol. 40
2004, Vol. 40文章信息
- 张飞萍, 陈清林, 尤民生, 侯有明.
- Zhang Feiping, Chen Qinglin, You Minsheng, Hou Youming.
- 毛竹林经营干扰、林下植物与冠层螨类之间的关系
- Relation between Management Disturbance and Undergrowth and Canopy Mites in Phyllostachys pubescens Forests
- 林业科学, 2004, 40(5): 143-150.
- Scientia Silvae Sinicae, 2004, 40(5): 143-150.
-
文章历史
- 收稿日期:2002-11-04
-
作者相关文章


2. 福建三明市三元区林业局 三明 365000;
3. 福建农林大学植物保护学院 福州 350002
2. Forestry Bureau of Sanyuan Sanming 365000;
3. College of Plant Protection, Fujian Agriculture and Forestry University Fuzhou 350002
近些年来福建、浙江一带毛竹叶部害螨的大面积危害,已成为毛竹(Phyllostachys pubescens)林三大效益发挥的最大障碍之一。有关这些害螨的种类、分布、危害、生物学、生态学、综合治理措施及其天敌的研究,已有了许多报道(崔云崎,1989;刘怀等,1999;余华星等,1991;张艳璇,1997;1999b;Zhang et al., 1998;张飞萍,1999;2000;2001)。作为竹林普遍且长期存在的害螨,其成灾的机制十分复杂,但最主要的原因是生境的改变,而气候的变化、人为的经营干扰是引起竹林生境改变的直接原因。清除毛竹林内的乔、灌木,将混交林改为纯林,是近些年主要的毛竹林低改措施(楼一平等,1997),现存的竹木混交林已为数不多。有研究认为,把混交林改为纯林是引起毛竹害螨大发生的原因之一(Zhang et al., 1999)。毛竹大面积纯林化已给人们在一定阶段内带来了诸多的经济效益。也有些林地出现地力衰退和产量下降的现象(楼一平等,1999),于是人们又加大了对竹林的耕作,施肥、垦复、劈山、喷山(对地被物喷施除草剂)已是除正常的伐竹和挖笋活动外最常见的经营方式,这些方式均表现为对毛竹林林下植物及土壤的干扰,但丧失植被保护的竹林,由于雨水的冲刷带走了腐殖质、表土和可溶性物质,使其地力衰退,也导致了毛竹林病虫害的发生(李正才等,2001)。本文通过对不同人为经营干扰下毛竹林林下植物群落和竹冠层螨类亚群落的研究,试图揭示经营干扰、林下植物、毛竹害螨三者之间的关系,其目的在于为毛竹害螨的生态控制乃至持续控制提供依据。
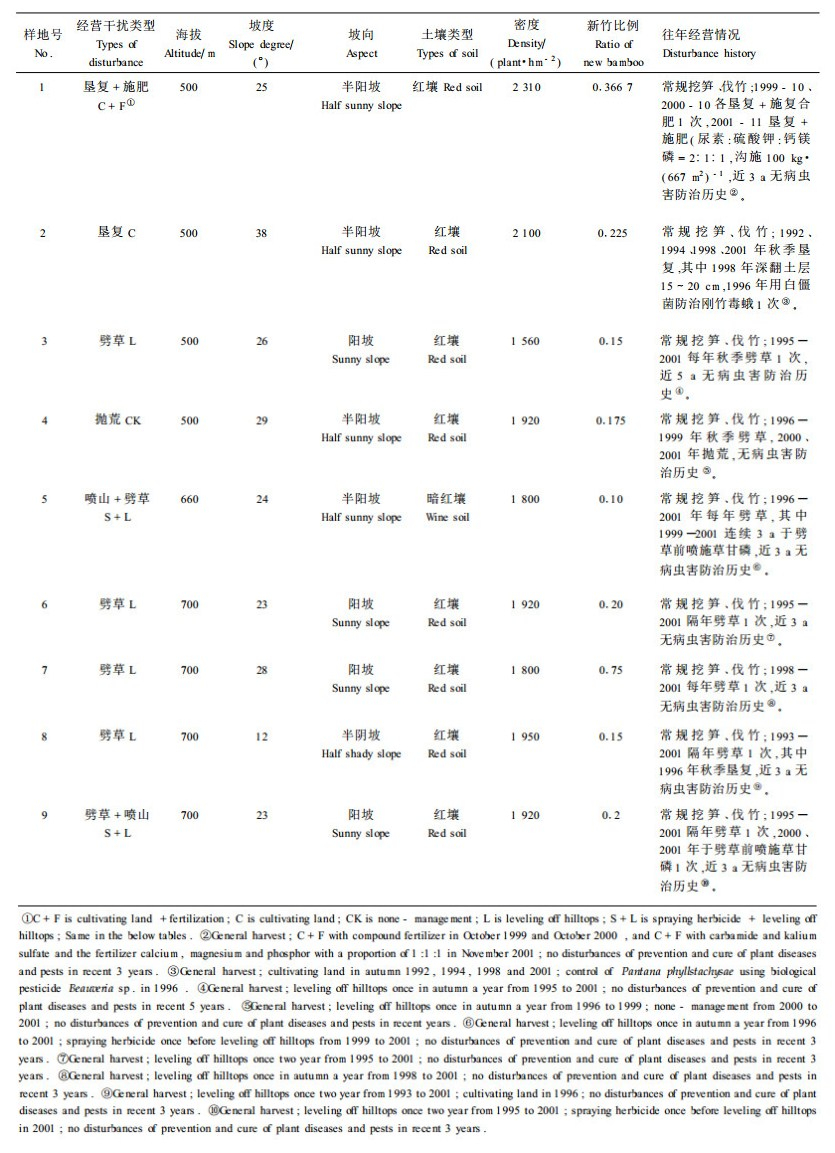
1 材料与方法 1.1 标准地概况及经营干扰类型的划分标准地位于福建省三明市三元区中村乡,该地属福建省毛竹中心产区,全乡现有竹林面积0.9万hm2,为全省乡镇有毛竹林地面积之首,其中约97%的毛竹林为纯林,当地居民80%的收入来自毛竹。2001年8月,通过样地踏查和户主家访,将毛竹纯林人工经营干扰划分为垦复施肥、垦复、劈草、喷山(喷施除草剂+劈草)以及抛荒(对照)5种类型,共设立9块标准地,各标准地面积约0.667 hm2,同年8—11月,对各标准地进行延续性经营干扰。标准地的基本情况及经营方式如表 1。
|
|
林下植物调查于2002年9月上述经营干扰实施前进行。对灌木层和藤本层的调查,在每标准地以平行线跳跃法设置5个10 m×1 m的长方形样方,详细记录各样方内木本、藤本植物的种类、数量、株高、盖度等信息,并将部分攀援状木本植物归入藤本层进行分析;对于草本层的调查,在上述样方的两端各取1个1 m×1 m的方形小样方,详细记录各小样方内草本植物的种类和盖度,苔藓植物归入草本层分析,各标准地共调查5个大样方和10个小样方(李正才等, 2001;林开敏,2001)。
1.3 毛竹林冠层螨类调查通过抽样技术的初步研究(另文发表),结合刘怀1)的方法,从2001年9月至2002年8月,每月中旬对各标准地进行抽样调查,具体方法为:各样地以五点取样法随机抽取20样株,用高枝剪于每样株冠层东、西、南、北、中5个方位各取2叶,装袋后带回室内镜检并记录叶片上的螨虫种类及数量,同时将网口直径为40 cm,深70 cm,用100目呢绒滤网制成的捕虫网捆绑于长竹杆上(呢绒滤网有利于取样时套住竹枝和从竹枝脱出),于每样株冠层中部随机抽取1枝条,套住后以均匀的力量连续抖动5次,每株调查2枝,将网中枯枝、落叶及螨虫等毒杀后袋装,带回室内镜检,统计、记录各种害螨及天敌捕食螨数。将剪叶调查结果和抖网调查的捕食螨数(剪叶调查不易捕获捕食螨)一并进行分析。
1) 刘怀.毛竹冠层节肢动物群落及竹育走螨生物学、生态学研究.[博士论文].北碚:西南农业大学,2001
1.4 物种多样性测定方法(赵志模等,1990;马克平等,1994;韩宝瑜等,2000;师光禄等,2002)利用以下指数进行分析:(1)Berger-Parker优势度指数d=Nmax/N;(2)相对丰盛度Pi=Ni/N;(3)丰富度指数S;(4)Shannon-Wiener指数H′=-Σ(PilgPi);(5)Simpson优势集中性指数C=ΣNi(Ni-1)/N(N-1);(6)Pielou均匀度指数J=-Σ(PilgPi)/lgS;(7)基于Gini指数的均匀度指数Jgi=〔1-Σ(PilgPi)〕/(1-1/S);(8)Gini多样性指数D1=1-ΣPi2;(9)多样性奇数测度D0=(ΣPi2)-1-1。
式中,S为物种数;N为群落中总个体数;Ni为第i种的种群数量,i=1, 2, 3, …,S;Nmax为群落中优势种群数量。选择式(1)~式(6)测定林下植物和竹冠层螨类的多样性,选择式(4)~式(9)计算二者多样性的相关系数。所有数据处理采用DPS或EXCEL软件包完成。
2 结果与分析 2.1 毛竹纯林林下植物种类组成及其多样性将林下植物分为灌木层、藤本层、草本层3个层次。1~9号样地的调查结果为,灌木层有36科,84种植物,其中物种数达4种以上的科有山茶科、蔷薇科、大戟科、樟科、桑科、紫金牛科、漆树科和冬青科,分别有物种8、8、6、5、5、4、4和4种,占该层总物种数的54.76%;相对丰盛度排于前10位的科有山茶科(0.199 7)、蔷薇科(0.151 5)、紫金牛科(0.099 8)、樟科(0.075 0)、鼠剌科(0.057 0)、禾本科(0.053 5)、杉科(0.051 7)、冬青科(0.042 6)、漆树科(0.031 8)和大戟科(0.030 3),这10科的个体数占灌木层总个体数的79.29%,其余26科仅占20.71%;相对丰盛度达0.01以上的种见表 2。
|
|

一般而言,相对丰盛度Pi≥0.1的种为优势种,0.05≤Pi<0.1之间为丰盛种,0.01≤Pi<0.05为常见种,0.001≤Pi<0.01为偶见种,Pi<0.001以下为稀少种或罕见种,可见林下植物中的优势科为山茶科和蔷薇科,丰盛种为毛竹小苗、白花悬勾子、细齿柃木、梨茶和杉木小苗。
藤本层有植物15科28种,物种数达到3种以上的科有菝葜科、蝶形花科、葡萄科和茜草科,分别有3、4、3和4种,占该层总种数的50%;相对丰盛度排于前5位的科为菝葜科(0.195 0)、茜草科(0.178 3)、夹竹桃科(0.150 4)、蝶形花科(0.128 2)和野牡丹科(0.097 5),占该层总个体数的74.94%,其中前4科为群落中优势科;相对丰盛度达到0.01以上的种见表 2,由表中可知,藤本层的优势种为光叶菝葜、紫花络石、玉叶金花,丰盛种为光叶菝葜、铁线莲、地NFDA5。草本层有植物26科30种,其中禾本科3种,茜草科和萝NFDA6科各2种,其余各科均只有1种;相对丰盛度达0.01以上的种见表 2,由表 2可知,草本层的优势种为金毛耳草、马唐草、提灯藓和铁芒萁,占草本层总盖度的77.52%;丰盛种为黑莎草、翠云草和五节芒,占总盖度的20.88%。
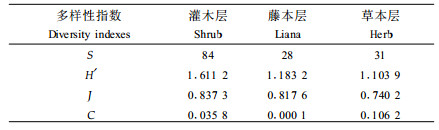
林下植物各层次的物种多样性指数见表 3,由表可知,各层次物种多样性和均匀度的大小序列均依次为灌木层>藤本层>草本层,物种丰富度为灌木层>草本层>藤本层,优势集中性为草本层>灌木层>藤本层。
|
|
为了减少空间异质性给比较分析带来的误差,本文选择空间距离较近的1~5号样地的调查资料进行分析,各样地的经营干扰情况见表 1,各样地的主要种类组成、多样性、均匀度状况分别见表 4、5。从表 4可知,各种类型干扰下,林下植物各层次的种类组成及数量分布格局存在一定差异,这与人为干扰方式、物种生长习性以及空间环境的异质性有关,如毛竹小苗在各种干扰方式下均存在,但在垦复施肥样地中,其相对丰盛度达到0.23,而其余大多数样地的相对丰盛度均在0.1之下,可以认为这是施肥引起竹鞭大量抽生竹芽(笋),加上垦复减少其营养及空间竞争所致;在种类组成方面以灌木层的差异最大,藤本层的差异次之,草本层差异最小;从各样地的优势度指数看,与对照(抛荒)相比,各种类型干扰均提高了灌木层的优势物种优势度,降低了藤本层和草本层的优势物种优势度。
|
|

|
|

由表 5可知,灌木层以抛荒和劈草的物种丰富度S最大,均为23,垦复施肥和垦复的次之,均为16,喷山+劈草的最小,仅为10;从其它指数看,与对照相比,受干扰样地林下植物的H′和J值均较低,C则较高, 可见各类经营干扰都降低了灌木层植物的多样性和均匀度,提高了其优势集中性。藤本层中从丰富度S看,劈草干扰和对照较接近,而其余3种的S较低;从H′和C看,劈草干扰的值比抛荒(对照)大,而垦复、垦复施肥与对照接近,仅喷山+劈草比对照低,因此可以认为,垦复、垦复施肥干扰降低了藤本层的物种丰富度,但对其物种的数量分布格局影响不大,而劈草干扰对物种的丰富度S影响不大,但对物种的数量分布格局影响较大,喷山+劈草干扰既降低了物种丰富度,又降低了物种分布的均匀度。草本层中除C值外,其余指数均比对照高,可见人为干扰可增加该层的物种丰富度、多样性和均匀度,这是由于不同干扰均减少了灌木层、藤本层对草本层植物的空间和营养的竞争,而草本层属于林下植物最低层,其竞争能力比其余2个层次低,藤本层属第2层,其竞争能力又比灌木层低,故对照的多样性及均匀性指数以草本层最低,藤本层次之,灌木层最高,这与2.1中分析结果一致。
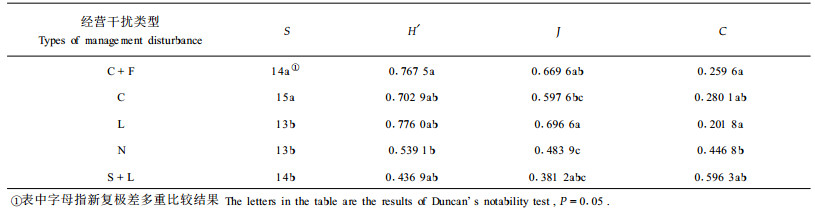
2.3 经营干扰下竹冠层螨类组成及其多样性同2.2仍然选择空间距离接近的1~5号样地数据进行分析,根据调查结果,竹冠层主要有螨类15种,其中相对丰盛度较大的害螨有竹刺瘿螨(Aculus bambusae)、竹缺爪螨(Aponychys corpuzae)、竹小爪螨(Oligonychus urama)、南京裂爪螨(Schizotetranychus nanjingensis),苔螨(Bryobia sp.);捕食螨有竹盲走螨(Typhlodrinus bambusa)、中国植绥螨(Phytoseius chinensis)和颈盲走螨(T.cervix)。各样地的物种多样性及均匀度指数见表 6,各类型干扰下的冠层螨类主要种及其相对丰盛度Pi见图 1。从表 6可知,各类经营干扰下竹冠层螨类物种丰富度指数S以垦复施肥和垦复样地显著高于对照和其它样地,物种多样性以垦复施肥显著高于对照,均匀度以垦复施肥和劈草经营显著高于对照,优势集中性则以垦复施肥和劈草样地显著低于对照,可见垦复施肥可显著提高竹冠层螨类的物种多样性,垦复施肥和劈草可显著提高其均匀度,降低其优势集中性。

|
图 1 不同干扰类型群落主要物种Pi值 Fig. 1 The Pi of main mite species in different communities 1垦复+施肥Cultivating land+fertilization; 2垦复Cultivating land; 3劈草Leveling off hilltops; 4抛荒Non-management; 5喷山+劈草Spraying herbicide+Leveling off hilltops. |
|
|
从图 1可知,垦复施肥竹林的害螨类优势种群为竹缺爪螨和竹刺瘿螨,垦复竹林的为竹缺爪螨、竹刺瘿螨和南京裂爪螨,但2者均以竹刺瘿螨相对丰盛度最高;喷山+劈草竹林以竹缺爪螨、竹刺瘿螨和竹小爪螨为优势种群,以竹小爪螨相对丰盛度最高;劈草竹林以竹缺爪螨、竹刺瘿螨、南京裂爪螨、竹小爪螨和颈盲走螨为优势种群,对照群落以竹缺爪螨、竹刺瘿螨为优势种群,2者均以竹缺爪螨相对丰盛度最高。天敌类群,除劈草群落的颈盲走螨相对丰盛度达到0.1以上外,其它群落的捕食螨Pi值均较低,但垦复施肥、垦复群落的Pi大于喷山+劈草和对照群落。
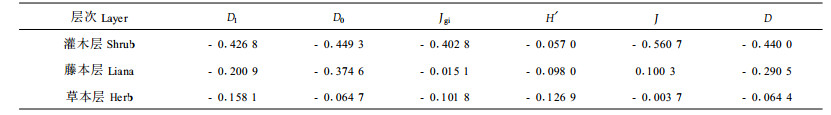
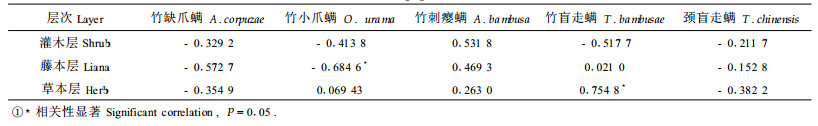
2.4 林林下植物与竹冠层螨类多样性之间的关系选择1~9号样地资料对林下植物各层次各多样性指数与竹冠层螨类相对应的多样性指数作相关分析,结果如表 7,从表可知,林下植物各层次对竹冠层螨类多样性的影响大小为灌木层>藤本层>草本层,但相关性均未达显著程度,仅灌木层接近显著水平。为了进一步考察林下植物与冠层螨类主要害螨、捕食螨种群的关系,选择林下植物各层次的Shannon-Wiener指数H′与1年中各阶段竹缺爪螨、竹刺瘿螨、竹小爪螨、竹盲走螨、颈盲走螨的平均每10叶螨口数量作相关分析,结果如表 8,从表可知,灌木层多样性与竹刺瘿螨呈正相关,与其余种群均呈负相关;藤本层多样性与竹刺瘿螨、竹盲走螨呈正相关,其余为负相关,其中与竹小爪螨呈显著负相关;草本层多样性与竹缺爪螨、颈盲走螨呈负相关,与其余种群呈正相关,其中与竹盲走螨种群数量呈显著正相关。综上可见,藤本层多样性显著不利于竹小爪螨种群的增长,草本层多样性显著地有利于竹盲走螨种群的增长。
|
|
|
|
随着毛竹经济地位的提高,人们对其经营管理日益精细,使毛竹林生产量在一定阶段内得到提高,但过度的经营干扰必然带来许多问题,如地力衰退、次要害虫大发生等。通过本研究发现,经营干扰对林下植物、冠层螨类多样性均产生了复杂的影响。一般情况下,人为经营干扰通过干扰林下植物以及寄主毛竹的生长情况而对冠层螨类产生间接影响,由于毛竹纯林内不具有乔木层,灌木层与竹冠层空间距离大,且林下植物与竹冠层螨类不具有直接的营养关系,故林下植物对冠层螨类种类组成及多样性的影响更可能通过改变林间的小气候和作为冠层螨类天敌或竞争物种的栖息地或营养资源库。从多样性的角度看,垦复施肥和劈草有利于提高竹冠层螨类的多样性或均匀性,结合两种经营方式下害螨的种群数量相对较低,因此这两种经营方式在生产中值得提倡。此外,经营干扰还可通过改变寄主毛竹的生长、生理生化功能等对冠层螨类产生影响,有关这方面的研究工作仍需进一步进行,才能深入地揭示经营方式与近些年毛竹害螨大发生之间的关系。
崔云琦. 1989. 我国为害竹林的叶螨. 昆虫知识, 26(5): 303-304. |
韩宝瑜, 张钟宁. 2000. 马尾松林直翅目昆虫群落的时空格局及其多样性和稳定性. 昆虫学报, 43(supp1): 143-150. |
楼一平, 盛炜彤, 萧江华. 1999. 我国毛竹林长期立地生产力研究问题评述. 林业科学研究, 12(2): 172-178. DOI:10.3321/j.issn:1001-1498.1999.02.010 |
楼一平, 吴良如, 李瑞成, 等. 1997. 竹林混交林改为毛竹纯林经营后的林分生长动态. 林业科学研究, 10(1): 35-41. DOI:10.3321/j.issn:1001-1498.1997.01.007 |
刘怀, 赵志模. 1999. 竹裂爪螨实验种群生命表研究. 西南农业大学学报, 21(6): 556-560. |
李正才, 李玉红. 2001. 两种不同类型毛竹林群落特征的初步研究. 竹子研究汇刊, 20(1): 45-49. DOI:10.3969/j.issn.1000-6567.2001.01.009 |
林开敏, 黄宝龙. 2001. 杉木人工林林下植物物种β多样性的研究. 生物多样性, 9(2): 157-161. DOI:10.3321/j.issn:1005-0094.2001.02.010 |
马克平, 刘玉明. 1994. 生物群落多样性的测度方法Iα多样性的测度方法(下). 生物多样性, 2(4): 231-239. DOI:10.3321/j.issn:1005-0094.1994.04.009 |
师光禄, 曹挥, 戈峰, 等. 2002. 不同枣园生态系统中昆虫群落及其多样性. 林业科学, 38(3): 94-101. |
余华星, 石纪茂. 1991. 南京裂爪螨的研究. 竹子研究汇刊, 10(2): 61-66. |
赵志模, 郭依泉. 1990. 群落生态学原理与方法. 重庆: 科学技术文献出版社重庆分社, 12.
|
张飞萍, 蔡秋锦, 钟景辉. 1999. 南京裂爪螨生物学及其发生与温度、降雨的关系. 福建林学院学报, 19(4): 372-374. DOI:10.3969/j.issn.1001-389X.1999.04.024 |
张飞萍, 陈玉鲁, 刘长士. 2000. 毛竹胸围与叶螨危害关联分析. 西南林学院学报, 20(3): 152-155. |
张飞萍, 蔡秋锦, 钟景辉, 等. 2001. 毛竹叶螨及其天敌捕食螨生态位的研究. 林业科学, 37(2): 56-61. |
张艳璇, 刘巧云, 林坚贞, 等. 1997a. 福建省毛竹叶螨种类, 分布及危害研究. 福建农科院学报, 12(3): 11-15. |
张艳璇, 刘巧云, 宋美官, 等. 1997b. 南京裂爪螨生活习性及防治研究. 植物保护, (5): 13-16. |
Zhang Yanxuan, Zhang Zhiqiang, Tong Xing. 1999. One cause of mite pest outbreaks in bamboo forests in Fujian, China: analyses of mite damage in monoculture versus polyculture forests. Systematic & Applied Acarolgy (Special publication), (4): 1-15. |
Zhang Yanxuan, Zhang Zhiqiang. 1998. Predation of Amblyseius longispinosus(Acari: Phytoseiidae) on Aponychus corpuzae(Acari: Tetrangkhidae). Systematic and Applied Acarolgy, (3): 53-58. |