 2004, Vol. 40
2004, Vol. 40文章信息
- 谢映平, 薛皎亮, 唐晓燕, 赵世林.
- Xie Yingping, Xue Jiaoliang, Tang Xiaoyan, Zhao Shilin.
- 绵粉蚧危害的花椒树对异色瓢虫的招引作用
- The Bunge Prickly-Ash Tree Damaged by a Mealybug, Phenacoccus azaleae Attracting the Ladybug, Harmonia axyridis
- 林业科学, 2004, 40(5): 116-122.
- Scientia Silvae Sinicae, 2004, 40(5): 116-122.
-
文章历史
- 收稿日期:2003-01-08
-
作者相关文章


天敌昆虫—植食性昆虫—寄主植物之间的食物链关系在很大程度上是靠三者之间的化学信息素来联系的,这在昆虫与植物,害虫与天敌的协同进化和害虫的生物防治领域具有重要意义。Vinson等(1981)、Turlings等(1990)、Dicke (1990)、Vet等(1992)、张瑛等(1998)研究了在植物—植食性昆虫—天敌昆虫三营养层次之间的化学信息素(semiochemicals or infochenmical)及其在害虫防治中的作用与利用途径,为害虫生态调控和生物防治指出了新的方向。我国在近10年来也做了不少工作,如娄永根等(1996)发现稻飞虱缨小蜂(Anagrus nilapavatae)对水稻品种挥发物的行为反应,并对虫害诱导的植物挥发物的基本特性、生态功能及释放机制和植物的诱导抗虫性作了研究报道。杜永均等(1994)比较详细地论述了植物挥发性次生物质在植食性昆虫、寄主植物和昆虫天敌关系中的作用机理。韩宝瑜等(2000)报道了七星瓢虫(Coccinella septempunctata)和异色瓢虫(Harmonia axyridis)对茶树和蚜虫的气味表现出行为反应。截止目前,研究记录的至少有13科24种植物与29种害虫和29种天敌昆虫之间表现出这种现象(娄永根等,2001)。
花椒绵粉蚧(Phenacoccus azaleae)属蚧总科(Coccoidea),粉蚧科(Pseudococcidae)的绵粉蚧属,原产日本,寄生杜鹃科植物。1988年首次在我国记录于内蒙古,寄生榆树(Ulmus pumila),虫口密度很少(汤枋德等,1988)。1994年发现该虫在山西省太行山区的花椒林发生危害,到1996年即从北部的盂县、五台,中部的左权县、黎城县,到南部的芮城县等地普遍发生,蔓延迅速,株虫口密度达到数万头,致使大面积花椒林受害。经调查,该蚧虫对与山西毗邻的河南、河北、陕西以及山东和甘肃等省的花椒林也造成危害。该虫1年1代,有两个危害高峰期,自3月下旬,出蛰的3龄雌若虫扩散到嫩芽和枝叶上取食,直到6月初,雌成虫产卵完毕,为第1个危害期,该期蚧虫取食量大,发育迅速,危害特别严重,常导致花椒树当年不能正常开花结果,产量骤减。7月下旬到越冬前,由第1和2龄若虫在叶片上吸食汁液危害,形成第2个危害期。由该虫连续危害2年,便会造成大花椒树死亡(Xie Yingping et al., 1997;1999)。
在林间已经发现,捕食绵粉蚧的多种天敌昆虫,如瓢虫、草蛉(Chysopa septempunctata)和蓟马(Aleurodothrips fasciapennis)等,其中异色瓢虫、蒙古光瓢虫[Exochomus(Anexochomus) mongol]、龟纹瓢虫(Propylaea japonica)是优势类群,其成虫和幼虫均捕食绵粉蚧,特别是异色瓢虫在春末夏初活动最盛,对绵粉蚧种群的控制作用显著(谢映平等,2002)。本文是在近年来本课题组对山西太行山花椒产区绵粉蚧发生与防治研究基础上,于1999—2001年又在盂县定点研究了受该蚧虫危害的花椒树(Zanthoxylum bungeanum)对天敌昆虫异色瓢虫的招引作用,并采用气相色谱-质谱联用(GC/MS)方法分析比较了受害和未受害花椒枝叶的化学成分,这对于掌握瓢虫与蚧虫及其寄主植物三者之间的化学信息,更好地开展生物防治,提供了新的资料。
1 材料和方法 1.1 实验林地概况固定实验林地选择在盂县北部花椒产区的梁家寨乡,第一类实验地在张家坪,面积20 hm2,属于山地梯田,花椒纯林。第二类实验地在梁家寨村的牛道坪林地,面积20 hm2,属于花椒林与粮食作物混交间种,花椒树成行排列。这两块林地在1995—1997年绵粉蚧发生都十分严重,一直作为本课题组研究花椒绵粉蚧的实验地。花椒树龄20 a左右,树高2~3 m,冠幅2.5~3 m。
1.2 试验材料的采集试验用瓢虫的采集 异色瓢虫对绵粉蚧的捕食作用主要在4月和5月份,在林间该瓢虫主要的色斑型有5种,即十八斑变型(H. axyridis form octadecade-punctata)、十九斑变型(H. axyridisvar. novemdeci mpunctata)、显现变型(H. axyridisvar. conspicua)、显明变型(H. axyridis var. spectabilis)和暗黄变型(H. axyridis form Yellow dark)。试验用瓢虫的虫态为成虫,雌性,一般为羽化后止产卵前的虫体,因为这个时期雌虫寿命长,为了产卵繁殖,需要寻找食物大量补充营养。在4—7月份进行。每次试验前在林地内采集足够量的5种色斑型成虫,分别放在500 mL空罐头瓶中,同时瓶中放入带蚧虫的枝条和叶片,带回实验室。
花椒枝梢的采集 实验前在林内采集新鲜标准枝梢,分为3种:带虫枝梢,即枝梢受蚧虫危害,试验时将蚧虫保留在枝梢上不取下。虫害枝梢,即采回带虫枝梢,试验时将蚧虫从枝梢上用镊子轻轻取下。正常枝梢,即未受蚧虫危害的枝梢。在林地将采集的新鲜枝梢立即放入冰壶,带回实验室,放入冰箱,作为试验材料。
1.3 花椒枝梢对异色瓢虫引诱活性的测定试验原理与测试盒的设计 实验原理是根据植物的化学防御体系,当植物受昆虫取食危害后,体内要产生生物化学反应,改变其挥发性物质的成分或强度,由此形成信号,可能引导天敌昆虫趋向受害的植物,捕食或寄生害虫。本实验的生物活性测试盒是对一个昆虫标本盒稍作改造,其长、宽、高分别为32、25、5.5 cm,盒盖为透明纱网,以便观察。在盒的中央放置一个硬纸片做成的直径为15 cm,高5 cm的圈,圈上均匀的留有向下的缺口,缺口高2 cm,宽2 cm,缺口之间间隔0.5 cm,测试过程中瓢虫可以在圈内外自由出入。试验用标本盒从未放过任何药物和昆虫,提前将盖打开,晾晒10余天,保证它和其中放置的硬纸片都没有影响昆虫行为的其他味道。
新鲜枝梢对异色瓢虫引诱活性的测试 将采回来的3种新鲜枝梢材料分别测试,每次取样量20 g,放在测试盒的中央纸圈内,将每次供试的异色瓢虫按色斑型分开测试,每次实验用10头活成虫,分为2组,其中5头放入圈内,另5头放在圈外,立即用纱网盖住,开始观察瓢虫或是趋向或是背离花椒枝梢的活动情况。3 min后,把盒子旋转180°,以消除光照和风向产生的影响。再过3 min后,记录进入圈内的瓢虫数量,每种枝梢材料对应各个瓢虫变型,重复测试6次,计算出瓢虫对试验的味源材料的趋向率。测定一般在白天9:00—18:00之间,晴天和室温条件下进行。
枝梢材料的馏分对异色瓢虫引诱活性的测试 每次用新鲜枝梢各20 g,分别用水浴蒸馏提取法,蒸馏时间30 min,收集馏分30 mL。测试时将馏分吸附于大约4 g的脱脂棉上,均匀放入圈内的塑料布上,然后接瓢虫试验,其余步骤与(2)相同。
1.4 受害枝梢和正常枝梢化学成分分析试验材料的预处理 将带回实验室的受害枝梢和正常枝梢各取6 g,分别剪碎,放入试管内,各加入15 mL二氯甲烷(CH2Cl2)。试管放入AS3120型超声波振动器中振动萃取30 min,共萃取2次。合并两次的提取液,过滤后自然浓缩至5 mL,再用吹送氮气法浓缩至2 mL,作为上机检测的样品。
试验仪器与样品检测 仪器采用气相色谱/质谱联用仪,英国ThermoQuest仪器公司生产,型号TraceGC2000/MS。检测时每次进样量1 μL。测试条件为:柱温初始50℃,初始时间1 min,第1次升温速率40℃·min-1,升至300℃,保持10 min;第2次升温速率10℃·min-1,升至310℃,保持10 min,最高温度控制为350℃。检测时仪器所用的毛细管柱均为30 mⅹ0.25 mm,内壁涂层厚度0.25 μm。
图谱解析与数据处理 经GC/MS检测结果获得气相色谱图和质谱图,利用该仪器上配备的有机物数据谱图库系统(Nist Library),对组分进行计算机自动检索,得到色谱图中各个峰对应的化合物、分子式、分子量和分子结构式。由计算机自动转换计算出色谱图上各个峰的面积和面积百分比。确定峰位,根据每个峰面积和总面积的比例,计算出各个组分的相对含量。
2 结果和分析 2.1 新鲜花椒枝梢对异色瓢虫的引诱活性试验发现,瓢虫被放入测试盒后,先向四周扩散,有趋光性的表现。绝大多数爬行,偶有欲飞行为。当稳定以后,瓢虫的触角向侧上方伸展摆动,下唇须不断地抖动,表现出对气味的探索行为。然后,瓢虫向不同的方向爬行,有的就穿过中央的圆圈上留的缺口,爬向盒的边缘,有的则进入缺口,一直在试验枝梢上慢慢来回爬行,有些就钻进枝梢丛中,象在寻觅食物。计数最后停留在圈内的瓢虫,与试验的总数相除,得到瓢虫对这种试验材料的趋向百分率。试验结果(见表 1)显示,异色瓢虫趋向受害枝条的百分率明显大于未受害的枝条,前者平均为57.0%和56.0%,后者为20.0%,差异极显著。说明枝条受害后增加了对瓢虫的引诱力;在试验中瓢虫对带有蚧虫的受害枝条的趋向率为57.0%,对不带蚧虫的受害枝条的趋向率为56.0%,二者几乎没有差异,说明花椒树受害后对瓢虫的引诱活性增加,主要是通过寄主植物挥发物质变化所致;在异色瓢虫不同的色斑型之间比较,发现十九斑变种对受害枝条的趋向率最高,为61.7% ~ 63.3%。与其它4种色斑型之间的差异极显著。其余3种色斑型即十八斑、显现、显明色斑型之间的差异不显著,其对受害枝条的趋向率在53.3% ~ 58.3%之间,暗黄色斑型的趋向率最低,为51.7% ~ 53.3%。说明即使是同一种瓢虫,它们的色斑型不同,在生物学方面还是有一些差异的。这为利用瓢虫科学防治蚧虫和深刻认识物种也具有意义。
|
|

本试验采用水浴蒸馏提取法,对花椒树枝条进行蒸馏,用收集的馏分作吸引异色瓢虫的试验,结果(表 2)显示,异色瓢虫对不取除蚧虫的受害枝梢馏分的趋向率为37.0%,对取除蚧虫的受害枝梢的趋向率为36.3%,二者之间没有显著差异。对未受害枝梢的馏分的趋向率只有10.3%,极显著的低于受害枝梢的馏分。说明蚧虫为害花椒枝条后,诱导其化学成分发生变化,使其对天敌昆虫异色瓢虫吸引力增加,而且这些物质通过水浴蒸馏的方法可以获得,仍然具有生物活性。在异色瓢虫的5种色斑型中,仍然以十九斑的色斑型对浸提液的趋向率高,而其他4种色斑型之间差异不显著。这一试验结果与采用新鲜受害枝梢的试验结果是一致的,起到了相互印证的作用。
|
|
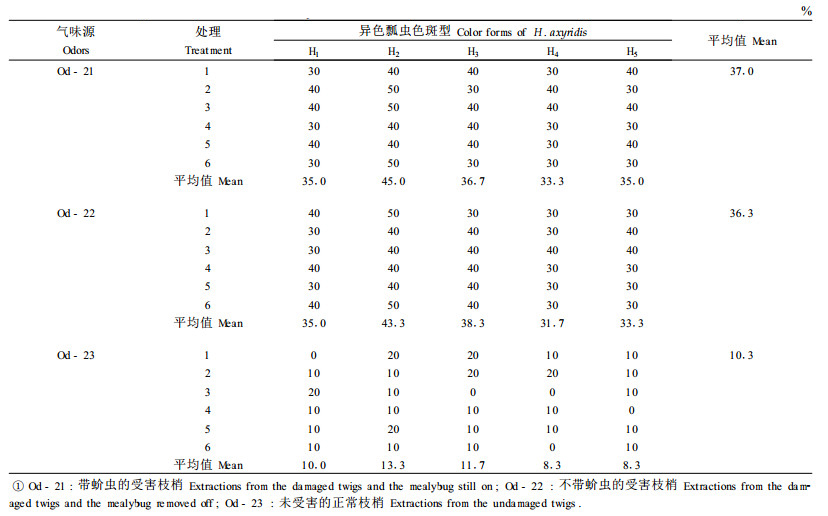
为了掌握受害枝梢的多少对异色瓢虫引诱力的变化,于2001年4月20日采用蚧虫危害的新鲜枝梢进行试验,新鲜枝梢的用量分别为10、20、30、40、50 g,结果见图 1。随着试验用虫害枝梢质量的增加,对异色瓢虫的引诱力增强,瓢虫表现出趋向率增高,这种规律对异色瓢虫的5种色斑型是一致的。同时,十九斑色斑型仍然表现出对引诱物的较高趋向率。排在第2位的是十八斑色斑型,然后是显现色斑型和显明色斑型,最后是暗黄色斑型。这一实验结果与前面的试验是吻合的。根据这个试验结果可以初步推断,较大面积的花椒林在受到绵粉蚧危害后,树体散发出的挥发性化学物质将会有一个群体效应,有利于招引更多的天敌昆虫向该林地趋集,发挥生物防治作用。

|
图 1 受害枝梢重量增加对异色瓢虫吸引力的影响 Fig. 1 The effect of the damaged twig's weight to the taxis of the ladybug, H. axyridis |
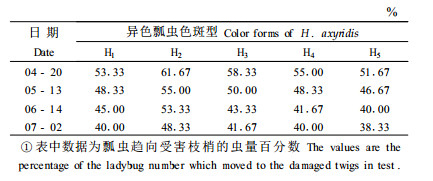
花椒绵粉蚧在3月中下旬开始出蛰,3龄雌性若虫从越冬的蜡茧内出来,扩散到嫩梢和嫩芽上,吸食汁液危害,发育2个龄期到达成虫,进入4月份为危害高峰期。4月下旬和5月上旬雌成虫开始分泌卵囊,并产卵于囊中,产卵完毕,雌成虫便死亡,不再危害。卵在囊中于6月份孵化,逗留囊中直到7月份才出来,扩散到嫩梢和叶片上,又开始取食危害,直到深秋。为此,本研究设计了从4月20日到7月4日期间的4次测试,每次均采用受害的新鲜枝条20 g,测试异色瓢虫对试验枝梢的趋向率。结果(表 3)显示,在蚧虫危害高峰期的4月20日测试,异色瓢虫对受害花椒枝梢的趋向率在51.67%~61.67%,在危害后期的5月13日,趋向率为46.67%~55.00%,绵粉蚧在6月份即早已停止危害,6月14日测试其趋向率为40.00%~53.33%,直到7月2日,其趋向率仍然达到38.33%~48.33%。说明花椒林受害后,树体内对天敌昆虫具有引诱活性的挥发性物质可以持续较长时间,这样,在受害的花椒林内及其周围,能形成持续较长时间的一个天敌昆虫引诱源,引诱天敌昆虫不断地向该林地趋集,并能逗留住,这样不仅在蚧虫开始危害时,而且在危害一段时间后,仍能诱导天敌昆虫来捕食蚧虫,发挥控制作用。
|
|
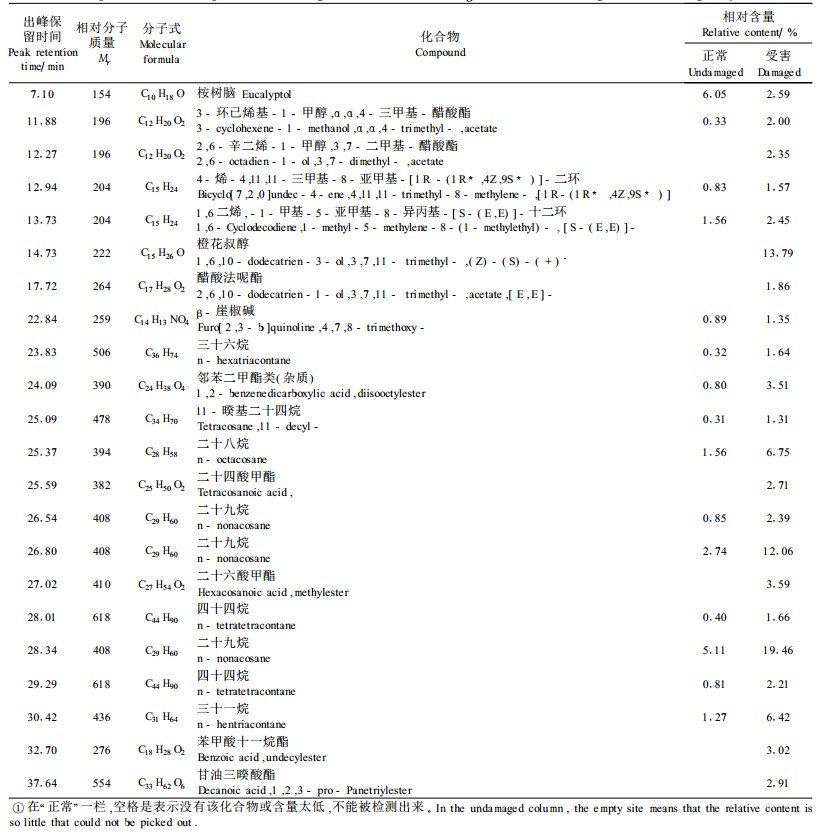
异色瓢虫对受害枝条的趋性明显高于正常的枝条,说明绵粉蚧危害诱导树体化学物质改变,在4月下旬采集受害和正常枝梢,提取其化学物质,通过GC/MS分析,比较二者化学组分的差异(表 4)。结果显示,在受害和正常枝梢中有15个峰位的化学组分相同,但受害枝梢的含量明显的高。其中3-环已烯基-1-甲醇, α, α, 4-三甲基-醋酸酯高出6倍,4-烯-4, 11, 11-三甲基-8-亚甲基-[1R -(1R*, 4Z, 9S*)]-二环高出2倍,几种直链烷烃的含量也增高3~6倍。在另7个峰位,只有受害枝梢出峰,说明这7个峰位显示的化合物在正常枝梢内含量极低,很难检测出来,或者根本就不存在,但经过蚧虫危害诱导,使枝叶内这种物质含量增加,容易检测出来,或者新产生形成这些化合物。这些化合物主要是高分子的长链或环状酯或醇。
|
|
本文研究证明,绵粉蚧危害后诱导花椒树体内的化学物质发生变化,释放出挥发性物质,对蚧虫的天敌昆虫异色瓢虫有明显的招引作用。这是害虫、寄主植物和天敌昆虫之间化学信息素联系的新例证,是生物协同进化的结果。这种招引作用的强度随着寄主植物受害数量和受害强度而增加,说明花椒受害,它的树木挥发物具有群体强化作用,有利于距离林分较远的天敌昆虫感受这种化学信息,从而向害虫发生的花椒林内聚集,这正是树木化学防御体系发挥的作用。试验还发现了这种招引作用在受害后能持续大约2个月的时间,这对于受害林分极为重要,它不仅可以将天敌昆虫招引来,而且可以使天敌昆虫在林内感受到持续的化学信息素的刺激,在林内停留的时间便延长,对害虫的控制作用增强。利用受害的新鲜枝梢及其水浴蒸馏收集的馏分招引异色瓢虫的实验结果相似,为人工利用这种化学信息素,开展花椒绵粉蚧的生物防治提供了重要的理论依据和实践途径。
用带蚧虫的花椒枝梢和取掉蚧虫的花椒枝梢试验对异色瓢虫的招引效果,二者差别很小,说明受害花椒树对天敌昆虫的招引主要是通过树木产生的化学物质来起化学通讯作用,蚧虫本身的挥发物质起的作用相对较小,这与Turlings等(1995)报道的植物受害后产生的挥发物质对天敌的引诱作用远远超过植食性昆虫本身或其遗留的粪便的结论是一致的。Dicke(1993)在研究其它树木受损伤后,发现其挥发性次生代谢物持续相当长一段时间,本研究结果证明在花椒树受绵粉蚧为害后也能持续释放挥发性化学信息素。
采用GC/MS联用技术,测试比较受害和未受害花椒枝梢内的化学物质变化,发现其中酯类物质明显增多。这与前人研究在苹果、山楂和甘蓝上发现的变化趋势相似(娄永根等,2001)。推测它们在诱导天敌昆虫过程中可能起重要作用。正如Pare等(1997a;1997b;1998)研究指出的那样,植食性昆虫危害除了导致植物释放绿叶性气味和增加植物体内一些储藏的挥发物组分或其前体转变成挥发物以外,还能促使植物合成并释放一些新的挥发组分,主要是萜烯类化合物。如他们在棉花上试验发现新合成的挥发物有一些非环状萜烯类化合物(E,E)-α-法尼烯,(E)-β-法尼烯,(E)-β-罗勒烯,芳樟醇,(E)-4,8-二甲基-1,3,7-壬三烯,(E,E0-4,8,12-三甲基-1,3,7,11-十三碳四烯以及吲哚是昆虫危害后新合成的。本次试验花椒受绵粉蚧危害后,枝梢内也有新合成的组分,与Pare等的结果具相似之处。但是,在虫害诱导反应中,植物产生的次生代谢物是复杂的,有些是增加某些组分,有些是减少某些组分,有些真正起引诱天敌昆虫的信息化合物的含量也可能是在微量级别上起作用。因此,对检测到的化学组分继续分离实验是必要的。前人的研究已经意识到天敌对植物源的反应可塑性较大,它在寻找寄主的过程中对植物气味容易产生学习行为,它可能把昆虫的信息源与植物的互益素联系起来(杜永均等,1994),异色瓢虫捕食的昆虫有蚜虫,蚧虫等同翅目昆虫,它对受害的花椒树的趋性在某种意义上就是将寻找蚧虫的目标与受害花椒树的气味结合的反应行为。这说明异色瓢虫具有学习特性,在生物防治中具有重要意义。
杜永均, 严福顺. 1994. 植物挥发性次生物质在植食性昆虫、寄主植物和昆虫天敌关系中的作用机理. 昆虫学报, 37(2): 233-250. DOI:10.3321/j.issn:0454-6296.1994.02.007 |
韩宝瑜. 2000. 异色瓢虫4变种成虫对茶和茶蚜气味行为反应. 应用生态学报, 11(3): 413-416. DOI:10.3321/j.issn:1001-9332.2000.03.021 |
娄永根, 程家安. 1996. 稻飞虱缨小蜂对水稻品种挥发物的行为反应. 华东昆虫学报, 5(1): 60-64. |
娄永根, 程家安.昆虫与植物相互关系的分子基础, 见: 程家安, 唐振华主编.昆虫分子科学.北京: 科学出版社, 2001, 139-170
|
汤枋德, 李杰. 1988. 内蒙古蚧害考察. 呼和浩特: 内蒙古大学出版社, 45-49.
|
谢映平, 赵世林, 唐晓燕.花椒绵粉蚧捕食性天敌的调查研究.见: 李典谟, 康乐, 吴钜文等.中国昆虫学会2002年学术年会论文集.北京: 中国科学技术出版社, 2002, 10: 496-501
|
张瑛, 严福顺. 1998. 虫害诱导的植物挥发性次生物质及其在植物防御害虫中的应用. 昆虫学报, 42(2): 204-213. |
Dick M, Sabelis M W, Takabayashi J, et al. 1990. Plant strategies of manipulating predator-prey interactions through allelochemicals: prospects for application in pest control. Journal of Chemical Ecology, 16: 3091-3117. DOI:10.1007/BF00979614 |
Dick M, Baarlen P V, Wessels R, et al. 1993. Herbivore induces systemic production of plant volatiles that attract predators of the herbivore: Extraction of endogenous elicitor. Journal of Chemical Ecology, 19(3): 581-599. DOI:10.1007/BF00994327 |
Pare P W, Tumlinson J H. 1997a. De novo biosynthesis of volatiles induced by insect herbivory in cotton plants. Plant Physiology, 114: 1161-1167. DOI:10.1104/pp.114.4.1161 |
Pare P W, Tumlinson J H. 1997b. Induced synthesis of plant volatiles. Nature, 385(2): 30-31. |
Pare P W, Tumlinson J H. 1998. Cotton volatiles synthesized and released distal to the site of insect damage. Phytochemistry, 47(4): 521-526. DOI:10.1016/S0031-9422(97)00442-1 |
Takabayashi J, Dicke M, Posthumus M A, et al. 1991. Variation in composition of predator-attracting allelochemicals emitted by herbivore infested plants: relative influence of plant and herbivore. Chemoecology, 2: 1-6. DOI:10.1007/BF01240659 |
Turlings T C J, Tumlinson J H, Lewis W J. 1990. Exploitation of herbivore induced plant odors by host-seeking parasitic wasps. Science, 250: 1251-1253. DOI:10.1126/science.250.4985.1251 |
Turlings T C J, Loughrin J H, Mccall P J, et al. 1995. How caterpillar-damaged plants protect themselves by attracting parasitic wasps. Proceedings National Academic Sciences of USA, 92(10): 4169-4174. DOI:10.1073/pnas.92.10.4169 |
Vet L E M, Dicke M. 1992. Ecology of info-chemicals use by natural enemies in a tritrophic context. Annual Review of Entomology, 37: 141-172. DOI:10.1146/annurev.en.37.010192.001041 |
Vinson S B Habitat location. In: Nordlund D A, Jones R, Lewis J eds. Semiochemicals: their role in pest control. New York: John Wiley & Sons, 1981: 51-68
|
Xie Yingping, Liu Hushan, Miao Zhenwang, et al. 1997. The outbreak of a mealybug, Phenacoccus azaleae Kuwana and its control in forest of bunge prickly ash. In: Chinese Academy of Forestry, USDA Forest Service, Resource Technology Institute eds. Resource Technology International Symposium Proceedings. Beijing: China Forestry Publishing House, 420-423.
|
Xie Yingping, Zhao Jinlong, Guo Yongping, et al. 1999. The biology of Phenacoccus azaleae, a pest of bunge prickly ash forest in northern China. Entomologica, 33: 377-382. |