 2004, Vol. 40
2004, Vol. 40文章信息
- 宋丛文, 包满珠.
- Song Congwen, Bao Manzhu.
- 天然珙桐群体的RAPD标记遗传多样性研究
- Study on Genetic Diversity of RAPD Mark for Natural Davidia involucrata Population
- 林业科学, 2004, 40(4): 75-79.
- Scientia Silvae Sinicae, 2004, 40(4): 75-79.
-
文章历史
- 收稿日期:2003-12-19
-
作者相关文章



RAPD技术作为一种简便、快速、易行的分子标记技术,近年来被广泛地应用于木本植物的遗传多样性和遗传结构的研究中。已经对辽东栎(Quercus liaotungensis)(恽锐等,1998)、大青杨(Populus ussuriensis)(苏晓华等,1997)、美洲山杨(Populus tremuloides)(Yeh et al., 1995)、椴树(Tilia)(Bielawski et al., 1997)、蒙古栎(Quercus mongolica)(夏铭等,2001)、马褂木(Liriodendron chinense)(李建民等,2002)等树种进行了报道。
珙桐(Davidia involucrata)是我国特有的珙桐科(Nyssaceae)单型属植物,其起源古老,是第三纪热带植物区系的孑遗种,被列为国家一级保护植物,也是世界著名的孑遗活化石观赏木本花卉植物。仅分布在甘肃、陕西、四川、湖北、湖南、贵州、云南等省的40多个县,呈星散状分布,现有原生群落面积十分有限,约4 500 hm2(张清华等,2000)。因此研究珙桐的遗传多样性,有效保存和合理利用珙桐的种质资源具有重要意义。本文利用RAPD技术对分布于我国的5个典型天然珙桐种群的遗传多样性、种群内和种群间的遗传变异进行了研究。
1 材料和方法 1.1 试验材料在湖北宣恩、四川峨嵋山、贵州梵净山、湖北神农架、甘肃文县等5个地方选择具有连续分布特征的天然林分作为研究的对象群体。在全林分随机抽取30~50株珙桐(每2株珙桐株间距不少于50 m),分别单株采集10~15片叶片,置于加入了足量干燥剂的塑料袋中密封,带回实验室后置于-20℃的冰箱中保存供试。
1.2 实验方法 1.2.1 DNA提取5个群体共186份样品(湖北宜昌40份、四川峨嵋山41份、贵州梵净山37份、湖北神农架30份、甘肃文县38份),采用CTAB法稍作修改后提取DNA。然后,测定DNA浓度和纯度,并通过琼脂糖电泳检查DNA的完整性,-20℃保存备用。
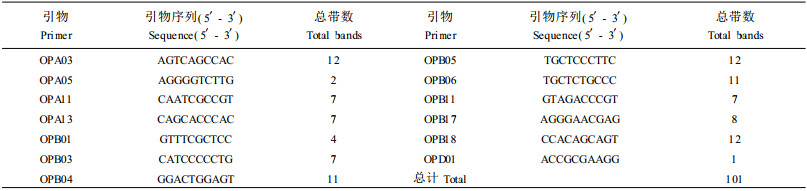
1.2.2 引物筛选随机引物采用Operon公司引物系列A、B、C、D共80个引物,经预备试验选择能产生多态性、清晰明亮的谱带,且重复性强的13个引物对所有个体DNA进行扩增。各引物名称、序列及检测带数见表 1。
|
|
RAPD扩增反应条件:94℃预变性3 min;然后进入40个循环,每个循环94℃变性1 min,45℃退火1 min,72℃延伸2 min;循环完成后于72℃维持7 min,最后于4℃保存。反应采用20 μL体系:H2O 11.6 μL,1×Buffer 2 μL,2 mmol MgCl2 0.8 μL,0.2 mmol dNTP 0.4 μL,0.4 μmol Primer 2 μL,1 Taq 0.2 μL,30 μg·μL-1 DNA 3 μL。扩增产物在1%含溴化乙锭的琼脂糖凝胶中电泳分离,最后在紫外灯下拍照记录结果(图 1)。

|
图 1 引物OPB05的RAPD扩增产物在神农架珙桐种群16个样品(S11—S26)中的分离 Fig. 1 Segregation of RAPD bands amplified with primer OPB05 In Shennongjia samples(S11—S26) of D. involucrata |
RAPD带按1/0标记,利用TFPGA(Tools for Population Genetic Analyses)软件(Version1. 31)进行数据分析。计算多态位点百分率、Nei's基因多样度、总的种群基因多样性、种群内基因多样性、种群间基因多样性、基因分化系数、基因流、种群间遗传同一性、种群间Nei' s遗传距离,并对5个种群进行UPGMA聚类分析。
2 结果与分析用13个随机引物对5个天然珙桐种群共186个个体的基因组DNA进行RAPD分析,共检测到101个可重复的位点,平均每个引物检测到的位点数为7.78。
2.1 多态位点百分率从表 2看出,在总计186个珙桐个体中共检测到总位点数为101个,其中多态位点98个,种内多态位点比率为97.03%。可以看出四川峨嵋山、贵州梵净山、湖北神农架3群体的多态位点的百分率明显高于湖北宣恩和甘肃文县群体。
|
|
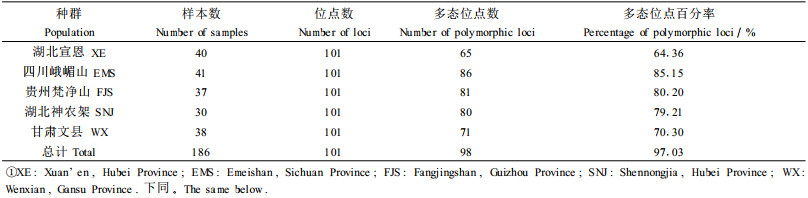
表 3表明了珙桐种内有着丰富的遗传多样性,基因多样度达到了0.333 6。各群体间遗传多样性有较大差异,贵州梵净山、湖北神农架和四川峨嵋山群体明显大于甘肃文县群体和湖北宣恩群体。
|
|

从表 4可以看到,珙桐种内平均遗传多样性为0.334 5,种群内平均遗传多样性为0.247 6,种群间的遗传多样性为0.086 9。基因分化系数达到了0.259 9,基因流为1.423 9,比较小。说明虽然珙桐大部分变异存在于种群内,但是由于珙桐受小种群效应,缺乏有效的基因流,致使种群间遗传变异占到了总变异的26%。
|
|

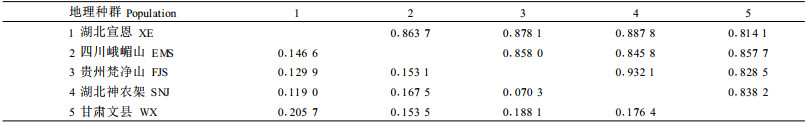
通过计算群体内个体间和群体间RAPD片段共享度,可以得到群体间的遗传同一度及Nei's遗传距离(表 5)。由表 5看出,珙桐群体间的遗传同一性以贵州梵净山和湖北神农架2个群体最大(0.932 1),其次是湖北宣恩与湖北神农架2个群体(0.887 8),湖北宣恩与甘肃文县的遗传同一性最小(0.814 1)。相反地,群体间的遗传距离则以湖北神农架与贵州梵净山最小(0.072 3), 其次是湖北宣恩与湖北神农架(0.119 0),以湖北宣恩与甘肃文县的遗传距离最大(0.205 7)。
|
|
根据表 5所列Nei's遗传距离, 使用UPGMA聚类法得到图 2所示聚类树形图。图 2聚类树形图显示,可以将5个珙桐天然群体分为2大类群体,即湖北宣恩、湖北神农架和贵州梵净山为一类,四川峨嵋山与甘肃文县为一类。据此,可以将珙桐划分为东南部和西北部2大种源区。

|
图 2 5个地理种源UPGMA聚类法得出的树状图 Fig. 2 Dendrogram showing Nei's genetic distance among 5 populations using UPGMA cluster analysis |
从每个群体中,按10、20、30个个体3个级别进行随机抽样,代表该群体进行遗传学计算,每个级别随机重复抽样10次。用POPGENE进行计算,所得结果见图 3~6。由图 3、4可见,有效等位基因数(Ne)和基因多样度(h)受取样个体数的影响很小, 即使在每个群体只取10个样时,也能较好地反映整个群体的状况,不过随着样本数的增大,这种波动逐渐减小,标准差逐渐减小,当取样数达到30个时,重复10次的值基本聚成一个点。而群体间的分化系数Gst和基因流Nm值的估算则明显受取样个体数的影响较大。就Gst而言,当每个群体的取样个体数为10个样本时,Gst值每次相差较大,且均偏离实际值较远;当每个群体取20个样本时,Gst相差显著减小;当样本数达30个时,Gst基本上能代表整个群体的水平(图 5)。对于基因流来说,则随着抽取个体数的增加,Nm值逐渐增大(图 6)。说明抽样样本数达30个时, 基本上能代表整个群体的遗传多样性水平。

|
图 3 有效等位基因(Ne)数和取样个体数之间的关系 Fig. 3 The value of effective number of alleles (Ne) obtained by the different number of sampled individuals (10×10, 20×10, 30×10) |

|
图 4 基因多样度(h)和取样个体数之间的关系 Fig. 4 The value of genetic diversity (h) obtained by the different number of sampled individuals (10×10, 20×10, 30×10) |
图中水平线为根据全部采样个体所得到的数值,每个小点为每次抽样所得到的遗传参数值。The horizontal line is based on all the D.involucrata individuals and each dot is generated from each sampling.下同。The same below.

|
图 5 基因分化系数(Gst)和取样个体数之间的关系 Fig. 5 The value of gene differentiation (Gst) obtained by the different number of sampled individuals (10×10, 20×10, 30×10) |

|
图 6 基因流(Nm)和取样个体数之间的关系 Fig. 6 The value of gene flow (Nm) obtained by the different number of sampled individuals (10×10, 20×10, 30×10) |
利用13个引物对5个珙桐天然群体进行了RAPD研究,结果表明,各群体的多态位点比率介于64.36%~85.15%。在对蒙古栎4个天然群体的DNA多态分析中,多态位点百分率为52.80%~66.45%(夏铭等,2001)。在对马褂木地理种群的DNA多态分析中,多态位点百分率在18.8%~36.2%(李建民等,2002)。从以上的对比中可以看出珙桐种内的DNA多态水平较高。Nei's基因多样性分析结果表明,珙桐群体内遗传多样性介于0.198 9~0.279 5,种内总的遗传多样性为0.333 6。高于一般针阔叶树种0.206的估算值(葛颂,1988),表明珙桐种内具有丰富的遗传多样性。从珙桐群体的多态位点百分率和遗传多样性来看,湖北神农架、贵州梵净山、四川峨嵋山3个群体的遗传多样性水平明显高于湖北宣恩和甘肃文县2个群体。
从珙桐遗传分化分析结果看,珙桐群体间遗传分化巨大,26%的遗传变异存在于群体间,与我国濒危树种马褂木(李建民等,2002),银杉(汪小全等,1996)等相似,而与广布性的树种相异。这与珙桐呈星散状残遗和间断分布在狭小范围之内,受小种群效应,以及缺乏有效的基因流(Nm=1.423 9)有关。
根据珙桐群体的遗传距离及其UPGMA聚类分析结果可以将珙桐划分为东南部和西北部2个种源区,东南部种源的遗传多样性略高于西北部种源。在进行原地保存时,全部保存当然是最好的,如果不能全部保存则可选择贵州梵净山、湖北神农架、四川峨嵋山3个群体进行保存,每个群体保存30个以上有效个体。在进行异地保存时,也可抽取这3个群体,每个群体抽取30个以上有效个体,则可有效保存珙桐的遗传多样性。
葛颂. 1988. 同功酶与林木群体遗传变异研究. 南京林业大学学报, 12(1): 68-77. |
李建民, 周志春, 吴开云, 等. 2002. RAPD标记研究马褂木地理种群的遗传分化. 林业科学, 38(4): 61-66. DOI:10.3321/j.issn:1001-7488.2002.04.010 |
苏晓华, 张绮纹, 郑先武, 等. 1997. 利用RAPD分析大青杨天然群体的遗传结构. 林业科学, 33(6): 504-511. DOI:10.3321/j.issn:1001-7488.1997.06.004 |
汪小全, 邹喻苹, 张大明. 1996. 松科系统发育的分子生物学证据. 植物分类学报, 35(2): 97-106. |
夏铭, 周晓峰, 赵士洞. 2001. 天然蒙古栎遗传多样性的RAPD分析. 林业科学, 37(5): 126-133. DOI:10.3321/j.issn:1001-7488.2001.05.022 |
恽锐, 钟敏, 王洪新. 1998. 北京东灵山辽东栎群体DNA多样性研究. 植物学报, 4(2): 169-175. DOI:10.3321/j.issn:1672-9072.1998.02.013 |
张清华, 郭泉水, 徐德应. 2000. 气候变化对我国珍稀濒危树种—珙桐地理分布的影响研究. 林业科学, 36(2): 47-52. DOI:10.3321/j.issn:1001-7488.2000.02.008 |
Bielawski J P, Pumo D E. 1997. Random amplified polymorphic DNA(RAPD)analysis of Atlantic coast striped kass. Heredity, 78: 32-40. DOI:10.1038/hdy.1997.4 |
Nei M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583-590. |
Yeh F C, Chang D K X, Yang R C. 1995. RAPD variation within and among natural populations of trembling aspen from Alberta. The Journal of Heredity, 86(6): 454-459. DOI:10.1093/oxfordjournals.jhered.a111620 |