 2004, Vol. 40
2004, Vol. 40文章信息
- 曹文强, 韩海荣, 马钦彦, 康峰峰, 蔺琛.
- Cao Wenqiang, Han Hairong, Ma Qinyan, Kang Fengfeng, Lin Chen.
- 山西太岳山辽东栎夏季树干液流通量研究
- Sap Flow Flux of Quercus liaotungensis in Summer in Deciduous Broad-leaf Forest of Taiyue Mountain in Shanxi Province
- 林业科学, 2004, 40(2): 174-177.
- Scientia Silvae Sinicae, 2004, 40(2): 174-177.
-
文章历史
- 收稿日期:2002-11-22
-
作者相关文章



树干液流指蒸腾在植物体内引起的上升液流。土壤液态水进入根系后, 由茎输导组织向上运送达冠层, 经由气孔蒸腾(包括角质层及皮孔蒸腾)转化为气态水扩散到大气中去。根部吸收的水分有99.8%以上消耗在蒸腾上(王沙生等, 1991)。通过精确测算茎流量, 可以基本确定植株蒸腾失水量。植物茎流测定对于建立土壤-植物-大气连续体模型, 以及探讨区域性水分供需规律, 验证抑蒸节水措施等都具有重要意义。
树干液流的测定方法很多,较为常用的有重量法及热量法等。重量法使用较早,一般认为可靠性低(Rutter, 1966;刘奉觉等,1997)。热量法测试茎流可根据测量原理分为2大类别:热脉冲法(heat pulse velocity)和热平衡法(thermal heat balance)。热脉冲法应用较广,目前绝大多数植物茎液流的测定都是基于热脉冲技术,但该方法测定的树干液流存在灵敏度调整和测定精度的问题(Swanson et al., 1981;Cohen et al., 1981)。热平衡法的优点是直接给出液流量,尤其是由Grainer(1987)发展起来的TDM法(thermal dissipation method)测茎流具有精度高、无需校正等优点,并经实验证明可信度高,且与独立观测结果具有一致性(Grainer, 1987;Grainer et al., 1996)。在我国,热平衡法的应用尚较为少见,严昌荣等(1999)曾利用Dynamax茎流仪对北京山区辽东栎(Quercus liaotungensis)和核桃楸(Juglans mandshurica)进行测定,而TDM法的应用仅见于对马尾松(Pinus massoniana)和栓皮栎(Quercus variabilis)液流的测定(鲁小珍,2001)。本项研究采用了澳大利亚ICT公司生产的ICT-2000自动气象站,基于热平衡法原理的TDM法对山西太岳山辽东栎的树干液流进行研究。
1 材料与方法 1.1 研究地概况实验于2001-07-08—08-25在山西省太岳山森林生态系统定位研究站(112°10′E,36°38′N)辽东栎落叶阔叶天然次生林中进行,海拔1 600 m。该区属暖温带半干旱大陆性季风气候,年均气温8.6℃,1月均温-10.4℃,7月均温17.4℃;年均降水量662 mm,雨量集中在7、8、9三个月,占全年降雨量的60%以上;年均日照2 500~2 700 h。大起伏喀斯特侵蚀高中山,主要为石灰岩,褐土、棕壤。
主要乔木树种为辽东栎,间有少量棘皮桦(Betula dahurica)、油松(Pinus tabulaeformis)、华北落叶松(Larix principis-rupprechitii)和核桃楸等。在实验地上选取1株具有代表性的林分平均木,胸径为23.2 cm,高13.2 m,去皮直径为21.6 cm,心材半径8.4 cm。样木生长良好,无病虫害,无被挤压现象。
1.2 热扩散法测定树干液流的基本原理TDP(thermal dissipation probe)是Grainer于1985年开发出的一种探针式探测器, 刺入茎内的木质芯材并接通恒定的电流以测定芯材的导热率。上升液流的冷却作用等效于芯材导热率的增加。热扩散法需要以液流量等于0时的加热功率及温度变化为依据进行零值较正。TDP探头包括2个探针,将2个探针插入树干,上方探针持续加热,下方探针为环境探针,通过测量这2个固定距离的探针的温度之差,用Grainer经验公式:V=0.000 119 K1.231,可求得液流速度。式中,V(m·s-1)为茎流速度,变量K=(dTm-dT)/dT,dT为加热的上方探头与下方探头的温差,dTm为记录的最大温差。得知不同深度的液流速度V,可据此求出整树的液流通量Flow=A×V。式中,A为有茎流木质部面积。
由于木质部不同部位的液流速度不同,TDP探头测定的是木质部径向液流速度的平均值。本实验的TDP-80探头长80 mm,有a、b 2个探测点,分别距尖端1.5 cm和7 cm,可分别测得边材和心材部分液流值。
1.3 仪器的安装在所选定样木的胸高(1.3 m)处安装仪器,以避免近地处冷液流的影响。刮出4 cm×10 cm的矩形,应尽量避免损害活组织。钻孔并将TDP-80探头插入钻孔中,并确保探头与树干紧密接触。为了防止太阳辐射对探头的影响, 在安装好探头的外层包上3层铝箔。将探头与数据采集器(data-logger)相应的接口进行连接,接通电源(12V的铅酸电池)。
数据采集间隔为15 s, 每10 min进行平均值计算并记录下来。
将TDP探头接到距样木3 m的ICT-2000自动气象站上,自动气象站可同步监测样木微环境下的林中空气温度、空气湿度、树冠顶部太阳辐射、5~10 cm深处土壤含水量(MP406探针测定)、5 cm深处土壤温度、风速、降雨量等气象因子。以上环境因子探头与TDP探头通过同一数据采集器(ZENO3200)控制。
2 结果与分析 2.1 蒸腾耗水的估计值表 1是7、8月份辽东栎在不同天气状况下的液流测定及统计结果,其中晴天以云量小于0.1或云量虽大于0.1,但云层极稀为准;雨天以降水量超过0.1 mm为准;多云天气和阴天一并统计。根据已知的液流日变化的数值和所测树木的去皮直径(21.6 cm),可以测得样木每日累积的液流量,从而计算其总蒸腾量。可知这株胸径为23.2 cm的辽东栎于7、8月份的总平均单位面积液流通量为60.25 cm3·cm-2d-1,蒸腾总量为1 346.7 L。
|
|

从测定的13 d的数据中选取典型的晴天、阴天、雨天分别分析辽东栎树干液流的日进程。图 1中各点的取值都是1 h内的平均值。从图 1可知,形成层以下7 cm处的探测点b所反映的木质部深处即心材部分的液流的日变化在晴、阴、雨天起伏都比较小,昼夜变化不明显,始终保持较低的值,一般小于1 cm·h-1,液流很微弱。液流主要集中在形成层以下1.5 cm处探测点a所测的边材部分,液流速度的日变化比较大。以下主要分析形成层以下a点液流的日变化状况。

|
图 1 辽东栎晴、阴、雨天树干液流日变化 Fig. 1 Diurnal change of sap flow rate in a sunny day, a cloudy day and a rainy day |
晴天树干液流从早8:00开始迅速上升,上午10:00达到其峰值12.61 cm·h-1,10:00—16:00液流一直保持较高的值,到16:00开始迅速下降,到19:00以后的夜间降至在小于1 cm·h-1的较低值的范围内摆动,直到次日晨7:00液流启动并迅速上升。在此过程中可发现:午间的液流值并没有随辐射和温度的持续上升而上升,而是保持平稳的高值,这可能与气孔的午间关闭有关。
阴天树干液流在12:00之前一直保持小于1 cm·h-1的低值,12:00液流值开始迅速上升,一直升到14:00时达到最大值8.71 cm·h-1,之后迅速下降,到20:00降到1 cm·h-1以下,并保持低值。阴天液流变化是一个典型的单峰型曲线,没有午休,显然是因为气孔不曾因强辐射而关闭。
7月12日除9:00—14:00,其余时间皆有雨,日降雨量为57.2 mm。液流的3个日变化峰值分别出现在6:00, 12:00, 17:00,但一直保持较低的值,在小于3.5 cm·h-1的范围内振荡,液流升降的辐度小。降雨增大了空气湿度,叶片内外的蒸汽压梯度大大降低,降雨还促使叶片气孔关闭,因而对上升液流有巨大制约作用。
3种天气状况下夜间都时常会有微弱的上升液流,有可能是根压使然(Clark et al., 1957)。
2.3 液流变化与环境因子的关系太阳辐射与温度太阳辐射与温度本身存在着一定的相关性。在图 1中,晴天的太阳辐射及温度与树干液流有明显的相同的趋势,太阳辐射的变化趋势与液流变化趋势更具有一致性。晴天与阴天气象因子的差别主要体现在辐射以及由辐射的差异导致的温、湿度变化上。反映在对液流速度的影响上特别体现在2个方面:一方面是对于液流启动时间的影响,液流清晨启动的时间与辐射启动的时间基本一致;另一方面体现在对其峰值变化的影响,阴天低辐射和低温对应着阴天的低的峰值,而在晴天午间的持续的高辐射和高温则对应着其高峰值和持续的高值。
空气相对湿度无论在晴天还是阴天空气相对湿度与液流速度都呈现出比较明显的相反的趋势;而在雨天,空气相对湿度一直为1,对应于雨天的液流速度低值也可以反映空气相对湿度对于液流的抑制作用。
降雨从图 3可得知,降雨对于液流有巨大的制约作用,雨天的液流速度始终保持低值,7月20日降雨时间长,强度大,日降雨量达57.2 mm,林内的空气相对湿度达到100%,极大地降低了叶片气孔内外的蒸汽压梯度。可见降雨通过影响其它因子的变化而影响液流变化。

|
图 2 晴、阴天液流速度与环境因子 Fig. 2 Diurnal change of sap flow rate and its environmental factors in a sunny day and a cloudy day |

|
图 3 阴天液流速度与土壤相对含水量的日变化 Fig. 3 Diurnal change of sap flow rate and soil water content in a rainy day |
土壤含水量比起太阳辐射和空气温、湿度来,土壤含水量的变化幅度要小得多,也慢得多。在晴天或阴天时,土壤含水量变化很小,几乎为一恒定值,并不能成为影响液流变化的主要因素。但在雨天则变化较大,由图 3可知,雨天液流速度的变化与土壤含水量的变化具有相同的趋势,但流速的变化仅仅在低值范围内振动,可以解释为根部的主动吸水的功能对土壤含水量变化的响应。
多元线性回归模型在以上分析的基础上,对于不同天气类型的液流速度(Flow)以及影响其变化的环境因子总辐射(x1)、空气温度(x2)、空气湿度(x3)、土壤相对含水量(x4)、降雨量(x5)进行逐步线性回归分析,可得以下回归模型:
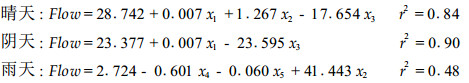
|
其中晴天和阴天的数据相关性较好,由回归分析的结果可知,晴天影响液流变化的主要因子为总辐射、空气温度和空气湿度,而阴天主要为总辐射和空气湿度。雨天的数据相关性较差,但可以得知土壤相对含水量对雨天液流的影响是最大的。
3 结论与讨论辽东栎样木7、8月份的总胸高截面平均单位面积液流通量为60.25 cm3·cm-2d-1,蒸腾总量为1 346.7 L。
辽东栎树干液流日变化在典型的晴、阴、雨天的不同天气状况下差异明显,在白天的液流启动时间、峰值及出现时间上都相差较大。晴天液流启动早,到达峰值的时间早,峰值大,且在达到峰值后液流值在午间保持较高水平。阴天为典型的单峰曲线,液流启动较晚,达到峰值后不久便迅速下降。雨天的液流变化规律性不强,起伏振荡多,且保持低值。7、8月份太岳山的天气类型多数情况下不是典型天气,以晴阴天气、阴雨天气组合较多,不稳定的天气因素增强了液流变化的不稳定性。
树干液流既由自身生理特性决定,同时与周围大气所发生的变化及土壤中水分的供应条件有着密切的关系。晴天和阴天影响液流变化的主要因子为太阳辐射、空气温度、空气湿度。雨天由于降雨造成了持续的低液流值,受土壤含水量影响较大。这些环境因子之间并不是独立存在和作用的,其间还存在着相互制约和相互协调。若想深刻理解各主要环境因子对树干液流的影响,仅仅依靠自然状态下的研究是不够的,必须根据控光、控水、控温及交叉作用的试验对各主要环境因子的依赖关系做数量分析,并在此基础上,逐步建立包括环境因子和树木生长参数的数学模型,去模拟树种的树干液流变化。
刘奉觉, 郑世锴, 巨关升, 等. 1997. 树木蒸腾耗水测算技术的比较研究. 林业科学, 33(2): 119-125. |
鲁小珍. 2001. 马尾松、栓皮栎生长盛期树干液流的研究. 安徽农业大学学报, 28(4): 401-404. DOI:10.3969/j.issn.1672-352X.2001.04.014 |
王沙生, 高荣孚, 吴贯明. 1991. 植物生理学. 第2版. 北京: 中国林业出版社, 192.
|
严昌荣, 陈兴国, 陈灵芝. 1999. 北京山区落叶阔叶林中核桃楸在生长中期的树干液流研究. 生态学报, 19(6): 794-797. |
Clark J, Gibbs R D. 1957. Studies on tree physiology. Ⅳ Further investigation of seasonal changes in moisture content of certain Canada forest tree. Can J Bot, 35: 219-253. DOI:10.1139/b57-021 |
Cohen Y M, Fuchs, Green G C. 1981. Improvement of the heat pulse method for determining sap flow in trees. Plant, Cell and Environment, 4: 391-397. DOI:10.1111/j.1365-3040.1981.tb02117.x |
Grainer A. 1987. Evaluation of transpiration in a Douglas fir stand by means of sap flow Measurement. Tree Physiology, 3: 309-320. DOI:10.1093/treephys/3.4.309 |
Grainer A, Huc R, Barigali S T. 1996. Transpiration of natural rain forest and its dependence on climatic factors. Agricultural and Forest Meteorology, 78: 19-29. DOI:10.1016/0168-1923(95)02252-X |
Rutter A J. 1966. Studies in the water relations of Pinus sylvestris in plantation, evaporation of intercepted water, and evaporation from the soil surface. J Ecol, 3: 393-405. |
Swanson R N, Whitfield D W A. 1981. Numerical analysis of heat pulse velocity theory. Journal of Experimental Botany, 32: 221-239. DOI:10.1093/jxb/32.1.221 |