 2004, Vol. 40
2004, Vol. 40文章信息
- 张真, 王军辉, 张建国, 张守攻.
- Zhang Zhen, Wang Junhui, Zhang Jianguo, Zhang Shougong.
- Bt转基因杨树对杨树昆虫群落结构的影响
- Effects of Transgenic Poplars to the Structure of Insect Community
- 林业科学, 2004, 40(2): 84-89.
- Scientia Silvae Sinicae, 2004, 40(2): 84-89.
-
文章历史
- 收稿日期:2002-08-29
-
作者相关文章



2. 中国林业科学研究院林业研究所 北京 100091
2. Research Institute of Forestry, CAF Beijing 100091
随着生物工程技术的迅速发展,分子遗传技术在农林有害生物管理中得到了广泛应用。将一些生物中的有用基因导入林木进行表达,成为保证林业生产的重要方法之一。杨树是我国尤其是北方地区防护林和用材林的重要树种,而杨树食叶害虫经常暴发成灾,给防护林建设带来极大的威胁。1991年中国林业科学院林业研究所同中国科学院微生物研究所在我国首次获得了欧洲黑杨(Populus nigra)转Bt毒蛋白基因植株,通过室内和大田抗虫性测定,对舞毒蛾(Lymantria dispar)和春尺蠖(Apochemia cinerarius)抗虫率达50%~100%(陈颖等,1995)。1994年在新疆北部地区建立欧洲黑杨Bt转基因实验林,1995—1997年试验区春尺蠖和梦尼夜蛾(Orthosia incerta)大暴发期间,显示出了明显的抗性(胡建军等,1999),但转基因实验林对环境的影响还没有进行过研究。
20世纪90年代以来,转基因生物的田间释放在世界各地不断进行,且释放规模逐年扩大,转基因生物的安全性受到学术界和整个社会的高度重视,转基因植物对非目标生物和生态系统的影响成为目前的一个研究热点。已有一些研究证明Bt转基因作物对非目标生物会产生不利影响,从室内和野外实验的结果说明Bt转基因玉米的花粉对深受人们喜爱的大斑蝶,又称君王斑蝶(Monarch Butterfly,Danaus plexippus)造成伤害(Losey等,1999;Jesse et al., 2000),尤其是Losey等(1999)在Nature上发表的论文,在世界各国引起很大的反响;Hilbeck(1998)等的研究表明转基因玉米对普通草蛉(Chrysoperla carnea)有一定的不利影响;崔金杰等(1999)研究说明种植转基因Bt抗虫棉对棉铃虫(Helicoverpa armigera)优势寄生性天敌齿唇姬蜂(Campoletis chlorideae)和侧沟绿茧蜂(Microplitis sp.)的寄生率、羽化率和蜂茧质量造成严重危害。Bt转基因植物在田间释放后可能产生的对有益昆虫和非目标种的毒性或不利影响是对环境影响的重要方面(Fitt,1995)。但也有实验得到影响不大甚至有利的结果,如夏敬源等(1999)的研究虽然表明Bt抗虫棉对棉铃虫寄生性天敌有严重危害,但却能有效地保护捕食性天敌,转基因棉田捕食性天敌总量较相应的对照明显增加。
Bt植物在自然界中释放的另一潜在风险是害虫的抗性进化,已有大量研究证明了鳞翅目的烟芽夜蛾(Heliothis virescens)、甜菜夜蛾(Spaloptera exigua)、海灰翅夜蛾(S.littoralis)、粉纹夜蛾(Trichoplusia ni)、小菜蛾(Plutella xylostella)、地中海粉螟(Anagasta kuehniella)、干果斑螟(Cadra cautella)、向日葵斑螟(Homoeosoma electellum)、印度谷螟(Plodia interpunctella)、纵色卷蛾(Choristoneura fumiferana),鞘翅目的黑杨叶甲(Chrysomela scripta)、马铃薯甲虫(Leptinctarsa decemilineata),双翅目的埃及伊蚊(Aedes aegypti)、五带色库蚊(Culex quinquefasciatus)、黄猩猩果蝇(Drosophila melanogaster)、家蝇(Musca domestica)在实验室条件下或田间对Bt产生了抗性(魏伟等,1999)。种植转Bt植物而导致害虫抗性的风险不能不引起重视,转Bt棉在美国南部的失败(Fox, 1996;Kaiser, 1996),进一步说明了问题的重要性。虽然害虫对Bt毒素的抗性是不可避免的,但是可以采取一系列有效的策略或对策来延迟抗性,常用的策略包括Bt作物的轮作、建立避难所、转入多个杀虫基因、高剂量方法、低剂量方法、Bt毒素的目标表达等(魏伟等,1999)。在林业上可以采取转基因和非转基因林木进行混交延迟抗性产生(卢孟柱等,1999)。
为了研究转基因杨树对非目标昆虫的影响及其害虫对Bt的抗性,分别种植了转基因纯林、转基因和非转基因1:1混交林,本文针对转基因Bt抗虫杨对杨树昆虫群落的影响进行了研究,结果报道如下。
1 研究地区概况和研究方法 1.1 林地概况研究地点主要有新疆玛纳斯平原林场和北京怀柔县北房镇。转基因杨树为欧洲黑杨转Bt毒蛋白基因植株。
玛纳斯平原林场位于新疆维吾尔自治区北部,地处天山北麓,准噶尔盆地南缘,43°13′N,86°37′E,海拔470 m,该地夏季炎热、干旱,冬季寒冷、多雪。林地土壤为沙壤土,适合许多杨树品种生长。1994年建立欧洲黑杨转基因实验林,面积约1 hm2,株行距3 m×4 m。共17个系号,其中包括14个转Bt基因欧洲黑杨系号、2个未转化的欧洲黑杨和健杨(P.euramericana(Dode)Guineir cv. ‘Robusta’)为对照。分布在6个区组中,每个区组的系号重复6次,单株小区,各系号的排列如图 1。实验林周围为本地的健杨。

|
图 1 转基因杨和非转基因对照的林间排列 Fig. 1 Distribution of Transgenic and Non-transgenic Clones in the Stand 图中编号为各系号的编号,CK2、CK3为未转换欧洲黑杨对照,本为本地健杨对照。 The numbers in the figure are number of the clones and CK2(P.nigra), CK3(P.nigra), 本(P.euramericana cv.`Robusta')are CK. |
怀柔北房镇,海拔9.7~64 m,土壤为沙壤质潮褐土,年平均气温9~12℃,年均降水650 mm,无霜期212 d,年平均日照2 700 h。北京转基因林于2001年春建立,一片为转基因纯林,位于宰相庄;一片为转基因和非转基因1:1混交林,位于小辛庄,非转基因杨为中林46(Populus×eur.cv.zhong1in 46),转基因和非转基因杨树采取隔行混交,面积均为6.7 hm2。
1.2 抽样方法新疆试验林的调查时间为2001年5月11日—13日,此时为新疆主要食叶害虫的危害期。北京怀柔试验林的调查时间为2001年9月12日,是杨扇舟蛾(Clostera anachoreta)的危害期。
调查时采用高枝剪套袋法进行抽样,套袋直径35 cm,长60 cm。调查时记录样枝各种昆虫的种类和数量。新疆试验林采取林冠层每个系号隔1株抽取1株进行调查,共抽样6株,每株分别抽取东、南、西、北不同部位的中下部枝条各1枝,共抽取转基因杨树12个系号72株288枝,对照24株96枝;在北京怀柔的转基因纯林和混交林分别采取对角线每行抽4株,分别抽取东、南、西、北不同方位的中部枝条,各样地共计抽取12株48枝。纯林和混交林分别调查叶片总数为1 008和913。
1.3 被害率统计在北京怀柔调查时将叶片被害程度分为全部吃光、3/4被吃光、1/2被吃光、1/4被吃光、1/8被吃光5个等级,危害等级值分别为1、0.75、0.5、0.25、0.125,被害率DR为:DR=[1/NΣ(NI×DI)]×100%,式中,N:调查叶片总数,NI:某一危害等级叶片总数,DI:危害等级值。
1.4 统计方法对被害率进行频率的差异显著性检验,对于转基因纯林与混交林之间的差异,nq > 30,采用u检验;混交林中转基因和非转基因杨树之间的差别,nq < 30,采用t检验,并进行连续性校正,可靠性均为95%(李春喜等,2000)。
1.5 群落特征分析对群落的多样性、均匀性和优势度指数进行分析,比较转基因和非转基因杨树、转基因纯林与混交林之间的昆虫群落的差异。其公式如下:
多样性指数H=-∑PilnPi,Pi为第i物种个体数占群落个体总数的比例;均匀性指数E=H/lns,s为物种数;优势度指数D=nmax/N,nmax为群落内ni的最大值,N为群落的个体总数。
2 结果与分析 2.1 虫口密度和被害率测定新疆转基因的各品系食叶害虫的平均虫口密度比对照低,但差异不明显,分别为每24枝2.6和3头,本地健杨对照最高,为3.5头。北京转基因纯林和混交林的叶片被害率分别为11.26%和18.48%,混交林大于纯林,但经频率的差异显著性检验,差异不显著(u=0.994 5,|u| < u0.05=1.96),且均在有虫不成灾的范围内。混交林内转基因和非转基因杨树的被害率分别为17.41%和19.54%,差异也不显著(t=-0.181 3,|t| < t0.05=2.014 7)。
2.2 群落组成结构新疆Bt转基因欧洲黑杨林主要的食叶害虫有梦尼夜蛾、春尺蠖;刺吸害虫主要有杨圆蚧(Quadraspidiotus gigas)和突笠圆盾蚧(Quadraspidiotus slavonicus)为群落的优势种;天敌昆虫主要有七星瓢虫(Coccinella odorata)、深点食螨瓢虫(Stethorus punctillum)、方斑瓢虫(Propylaea quatuordecimpunctata (Linnaeus))、菱斑巧瓢虫(Oenopia conglobata(Linnaeus))、二十二星瓢虫(Psyllobora vigintiduopunctata)。
北京欧洲黑杨林群落中的主要食叶害虫有杨扇舟蛾、杨叶蜂(Pristiphora benjingensis),天敌主要有龟纹瓢虫(Propylaea japonica)、异色瓢虫(Harmonia axyridis)。不同林分节肢动物群落结构见图 2-5。

|
图 2 转基因杨树昆虫和蜘蛛群落结构(新疆玛纳斯2001-05) Fig. 2 Insect and spider community structure of transgenic poplars (Malasi, Xinjiang May 2001) |

|
图 3 对照林昆虫和蜘蛛群落结构(新疆玛纳斯2001-05) Fig. 3 Insect and spider community structure of non-transgenic poplars (Malasi, Xinjiang May 2001) |

|
图 4 转基因纯林昆虫和蜘蛛群落种类及比例(北京怀柔2001-09) Fig. 4 Insect and spider community structure of pure trasgenic poplars (Huairou, Beijing September 2001) |

|
图 5 转基因混交林昆虫和节肢群落种类及比例(北京怀柔2001-09) Fig. 5 Insect and spider community structure of 1:1 mixed trasgenic poplars and non tragsgenic popars(Huairou, Beijing September 2001) |
新疆转基因和非转基因杨的群落结构相似,区别在于转基因杨树群落中的杨梦尼夜蛾、杨尺蠖等食叶害虫所占比例略小于对照,分别为2.4%和4%;瓢虫、草蛉、蚂蚁等捕食天敌所占比例明显比对照高,分别为46%和28%;刺吸害虫比对照低,分别为45%和62%。
从北京的调查结果看,转基因纯林与转基因混交林的群落结构发生了很大的变化,纯林中数量最大的是杨叶蜂,0.75头·枝-1,占58%:而混交林中数量最大的是杨扇舟蛾,1.39头·枝-1,占78%。由于Bt主要作用于鳞翅目昆虫,而对膜翅目影响不大,所以转基因纯林中杨叶蜂的密度较高,成为优势种。另一个明显的差别是转基因混交林中瓢虫的数量明显比转基因纯林高,分别为0.21头·枝-1和0.021头·枝-1,相差9倍。蜘蛛的数量是纯林中较多,分别为0.125头·枝-1和0.062 5头·枝-1,相差1倍。
2.3 群落的多样性和均匀性新疆的转基因和非转基因杨树及北京的转基因纯林和混交林的群落特征指数如表 1-2。表 1结果说明,新疆转基因和非转基因杨树群落各多样性指数差异不大。
|
|
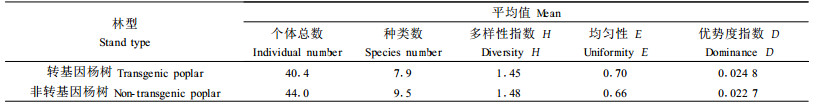
|
|
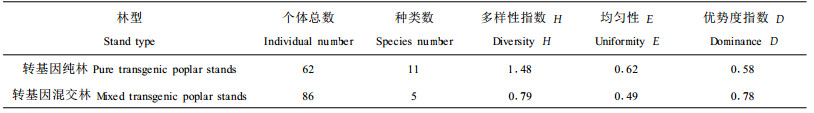
表 2结果表明纯林中昆虫个体总数较少,但多样性和均匀性较高,优势度指数较低。
3 结论和讨论新疆转基因杨树和非转基因杨树之间群落结构和多样性差别不大,而北京的转基因纯林和混交林之间无论从群落结构还是多样性都有较大的差异,这是由于新疆转基因林面积小,且各品系之间是相间排列,相互之间的影响较大,最终群落趋于一致;而北京的转基因林和转基因林与非转基因混交林林面积较大,其差异就明显显露出来。多大的规模下对害虫抑制作用最好,对非目标昆虫影响最小,是需要深入研究的课题。
Bt转基因杨树会改变其昆虫群落结构,由于Bt主要作用于鳞翅目昆虫,而对膜翅目影响不大,在抑制鳞翅目昆虫的同时,有利于其它食叶昆虫,所以转基因纯林中杨叶蜂为优势种,而在转基因与非转基因混交林中以杨扇舟蛾为优势种。这与Wilson等(1992)的研究相似。Wilson研究表明转Bt基因棉由于对鳞翅目害虫的抗性减轻了棉叶的危害,从而促进了其它害虫种群的增长,转Bt基因棉间接地促进了甘薯白粉虱(Bemisia tabaci)等的种群增长。中棉所29和中棉所30应用表明:棉蚜(Aphis gossypii)、棉叶蝉(Empoasca biguttula)、棉叶螨(Tetranychus cinnabarinus)、棉蓟马(Thrips tabaci)、棉盲蝽(Lygus sp.)、白粉虱(Trialeurodes vaporariorum)等刺吸式害虫的发生有加重趋势,可能是因为鳞翅目害虫减少或鳞翅目害虫的减少使得寄生性天敌减少而导致刺吸式害虫种群上升(崔金杰等,1999)。在转基因纯林中杨叶蜂能不能造成明显的经济损失?还需要进一步进行监测和研究。不同杨树种类对杨叶蜂的抗性是不一样的(周淑芷等,1995),为了避免其它非鳞翅目食叶害虫的大量危害,在转基因时应采用对非鳞翅目食叶害虫有抗性的种类或品系。
Bt转基因纯林的多样性和均匀性都较高,有利于系统的稳定性,但瓢虫数量明显较少,而新疆转基因林中转基因杨树群落的瓢虫比非转基杨树多。蜘蛛的数量是纯林中较多。所以Bt转基因杨树对天敌的影响还需进行详细研究。
营造转基因与非转基因林木的混交林是防止害虫对转基因林木产生抗性的途径之一(卢孟柱等,1999),这为敏感害虫群体设置了庇护所,通过保护敏感群体而限制了抗性个体的发展,不采用昆虫的防治措施,确保大量敏感性昆虫总是与附近的一些抗性昆虫交配,达到防止抗性群体产生的目的。本研究结果表明Bt转基因纯林抑制目标食叶害虫的作用较好,但由于有利于其它非鳞翅目害虫,总被害率与1:1的混交林之间差异不显著,后者是否足以使其长期有虫不成灾又能有效地防止抗性的产生?还需要进一步进行长期的监测和研究。
Bt转基因植物释放到环境后对生态系统的影响是多方面的,对其生长环境内的其它植物、微生物和各种动物都有可能产生影响,不仅会影响到目标害虫,也会直接地或间接地影响到其它非目标生物。这种影响的程度和对生态系统是否会造成危害,在大量种植之前必须从基因、基因组、个体、种群、生态系统各级水平上进行详细的研究,根据研究的结果作出风险评价并制定风险管理对策。Fitt(1995)总结了转基因棉花对环境的可能影响,从植物本身对环境影响的角度包括:1)杂草化;2)基因逃逸到临近植物;3)对有益昆虫和非目标昆虫的不利影响;4)棉花种子或枝叶对非目标种的毒性影响。从鳞翅目害虫和其它种对Bt棉的反应考虑可能有以下几方面的影响:1)目标害虫对Bt产生抗性;2)次要害虫对Bt产生抗性;3)由于减少化学农药,产生一些新的或次期害虫。针对Bt转基因杨树,除第一种危害的可能性外,其他影响都比较大,由于林木的多年生特点,对非目标生物的影响和产生抗性的可能性更大,尤其需要加强研究并制定相应的风险管理措施。本研究初步说明了Bt转基因杨树对杨树昆虫群落结构和非目标种的一些影响,说明了进行进一步深入的研究必要性和重要性。
陈颖, 韩一凡, 李玲, 等. 1995. 苏云金杆菌杀虫晶体蛋白基因转化美洲黑杨的研究. 林业科学, 31(2): 97-102. |
陈颖, 李强, 李玲, 等. 1996. 抗虫转基因欧洲黑杨的Western印迹法分析. 林业科学, 32(3): 274-276. |
崔金杰, 夏敬源. 1999. 转Bt基因棉对天敌种群动态的影响. 棉花学报, 11(2): 84-91. |
胡建军, 刘庆一, 王克胜, 等. 1999. 欧洲黑杨转Bt毒蛋白基因植株大田抗虫性测定. 林业科学研究, 12(2): 202-205. DOI:10.3321/j.issn:1001-1498.1999.02.015 |
李春喜, 王志和, 王文林. 2000. 生物统计学. 北京: 科学出版社.
|
李祎, 陈颖, 李玲, 等. 1996. 抗菌肽LcI基因转化杨树的阶段研究. 林业科学研究, 9(6): 646-649. DOI:10.3321/j.issn:1001-1498.1996.06.015 |
卢孟柱, 韩一凡, 杜生明. 1999. 林木基因工程风险评估和安全管理现状. 林业科学研究, 12(3): 325-331. DOI:10.3321/j.issn:1001-1498.1999.03.017 |
钱迎倩, 田彦, 魏伟. 1998. 转基因植物的生态风险评价. 植物生态学报, 22(4): 289-299. DOI:10.3321/j.issn:1005-264X.1998.04.001 |
钱迎倩, 魏伟, 桑卫国, 等. 2001. 转基因作物对生物多样性的影响. 生态学报, 21(3): 337-343. |
钱迎倩, 魏伟, 田彦, 等. 1999. 转基因作物在生产中的应用及某些潜在问题. 应用与环境生物学报, 5(4): 427-433. DOI:10.3321/j.issn:1006-687X.1999.04.019 |
田颖川, 李太元, 莽克强, 等. 1993. 抗虫转基因欧洲黑杨的培育. 生物工程学报, 9(4): 291-297. DOI:10.3321/j.issn:1000-3061.1993.04.017 |
王学聘, 韩一凡, 戴莲韵, 等. 1997. 抗虫转基因欧美杨的培育. 林业科学, 33(2): 69-74. |
魏伟, 钱迎倩, 马克平. 1999. 害虫对转基因Bt作物的抗性及其管理对策. 应用与环境生物学报, 5(2): 215-228. |
夏敬源, 崔金杰, 马丽华, 等. 1999. 转Bt基因抗虫棉在害虫综合治理中的作用研究. 棉花学报, 11(2): 57-64. |
周淑芷, 黄孝运, 张真, 等. 1995. 北京杨锉叶蜂研究. 林业科学研究, (5): 554-563. |
Fitt G P. 1995. Field evaluation of transgenic cotton in Australia:environmental considerations and consequences of expanding trial size. Biosafety Scientific Findings and the Need for a Protocol, An Independent Expert Report Prepared for the Biodiversity Convention: 37-47. |
Fox J L. 1996. Bt cotton infestations renew resistance concerns. Nat Biotech, 14: 1070. DOI:10.1038/nbt0996-1070 |
Hibeck A, Baumgartner M, Fried P M, et al. 1998. Eeffect of transgenic Bacillus thuringiensis corn fed prey on mortality and development time of immature Chrysoperla carnea(Naeuropter:Chrysopidae). Environ Entomol, 27(2): 480-487. DOI:10.1093/ee/27.2.480 |
Huang F, Higgins R A, Buschman L L. 1997. Baseline susceptibilty and changes insuscepibility to Bacillus thuringiensis subsp.Kursttaki under selection pressure in European Corn Borer(Lepidoptera:Pyralidae). J Econ Entomol, 90(5): 1137-1143. DOI:10.1093/jee/90.5.1137 |
Jesse L CH, Obrycki J J. 2001. Field deposition of Bt transgenic corn pollen:lethal effects on the monarch butterfly. Oecologia(Berl.), 125: 241-248. |
Kaiser J. 1996. Pest overwhelm Bt cotton crop. Science: 273, 423. |
Losey J E, Raynor L S, Carter M E, et al. 1999. Transgenic pollen harms monarch larvae. Nature: 399-214. |
Tabashnik B E. 1997. Seeking the root of insect resistance to transgenic plants. Proc Natl Acad Sci USA, 94: 3488-3490. DOI:10.1073/pnas.94.8.3488 |
Tabashnik B E, Cushing N L, Finson N, et al. 1990. Field development of resistance to Bacillus thuringiensis in Diamondbank Moth(Lepidoptera:Plutellidae). J Econ Entomol, 83: 1671-1676. DOI:10.1093/jee/83.5.1671 |
Tabashnik B E. 1994. Evolution of resistance to Bacillus thuringiensis. Annu Rev Entomol, 39: 47-79. DOI:10.1146/annurev.en.39.010194.000403 |
Wilson F D, Flint H M, Deaton W R, et al. 1992. Resistance of cotton lines containing a Bacillus thuringiensis toxin to Pink Bollworm and other insects. J Econ Entomol, 85(4): 1516-1521. DOI:10.1093/jee/85.4.1516 |