 2004, Vol. 40
2004, Vol. 40文章信息
- 喻方圆, 徐锡增, Guy Robert D.
- Yu Fangyuan, Xu Xizeng, Guy Robert D.
- 水分和热胁迫处理对4种针叶树苗木气体交换和水分利用效率的影响
- Effects of Water Plus Heat Stress on Gas Exchange and Water Use Efficiency in Four Species of Coniferous Seedlings
- 林业科学, 2004, 40(2): 38-44.
- Scientia Silvae Sinicae, 2004, 40(2): 38-44.
-
文章历史
- 收稿日期:2002-09-26
-
作者相关文章


2. 加拿大不列颠哥伦比亚大学 温哥华 V6T 1Z4
2. The University of British Columbia Vancouver Canada V6T 1Z4
杉木(Cunninghamia lanceolata)和马尾松(Pinus massoniana)是我国南方主要造林树种,每年造林育苗量约为10亿株。花旗松(Pseudotsuga menziesii)和北美乔柏(Thuja plicata)是北美的重要造林树种,每年的育苗量也很大。但在育苗过程中,这些苗木常会遭受各种环境胁迫,特别是夏季的高温和干旱胁迫,常常严重影响苗木生长,甚至造成苗木死亡。了解苗木对环境胁迫的生理反应,并在育苗过程中及时采取相应的技术措施,是培育高质量苗木的前提。最常用来反映苗木遭受环境胁迫程度的生理指标有叶绿素荧光的变化(variable chlorophyll fluorescence)、气体交换参数和可溶性糖含量等(Sinclair et al., 1985; Osório et al., 1994)。这些生理指标已成为植物生理生态领域研究的热点,相关报道很多。但目前涉及杉木和马尾松的报道却很少,特别是这2个树种对水分和热胁迫的生理反应鲜有报道。本文模拟杉木、马尾松苗木生长过程中可能遭受的高温和干旱胁迫,揭示高温和干旱条件下苗木适应环境胁迫的生理机制,为培育高质量的杉木和马尾松苗木提供依据。本研究同时选择花旗松和北美乔柏2个温带树种进行试验,一方面可以为亚热带树种杉木、马尾松提供一个参照,另一方面也能为这些温带树种对环境的适应性提供有价值的数据。
1 材料与方法 1.1 试验材料杉木和马尾松种子(均采自中国四川)于2000年10月26日播于D40型容器中。北美乔柏(种批号B6542,采自加拿大BC省)和内陆花旗松(种批号B4587,采自加拿大BC省)种子于2000年9月29日播于D40型容器(美国Stuewe & Sons公司)中。D40型容器深25.4cm, 直径6.35cm。播种量根据各树种的发芽率而定,每穴3~5粒种子。容器中的培养基为7:2:1 (v:v:v)的泥炭:蛭石:消毒矿物土壤。容器置于加拿大不列颠哥伦比亚大学(UBC)校园里的自动化温室内。温室的环境条件见表 1。
|
|
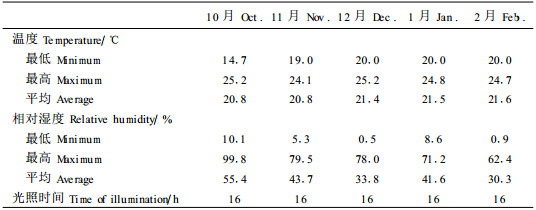
上述容器苗培育约5个月后,按设计移入4个人工气候箱(加拿大Conviron公司)中,并实施周期性水分和热胁迫处理,处理时间从3月2日到4月7日。4个人工气候箱中,2个温度设定为:头天下午15:00到次日上午9:00为28℃,当日上午9:00到当日下午15:00为35℃。另2个温度设定为:头天下午15:00到次日上午9:00为20℃,当日上午9:00到当日下午15:00为25℃。4个人工气候箱的光照强度均在280~330μmol·m-2s-1。每个树种在每个人工气候箱中有24~30株苗木,将这些苗木平均分成3组,分别施以不同的水分胁迫处理:干旱,每6d浇水一次;轻度干旱,每4d浇水一次;湿润,每2d浇水一次。浇水时均需将苗木浇透。总共进行了3个循环的干旱处理,每个循环12d,总计36d。
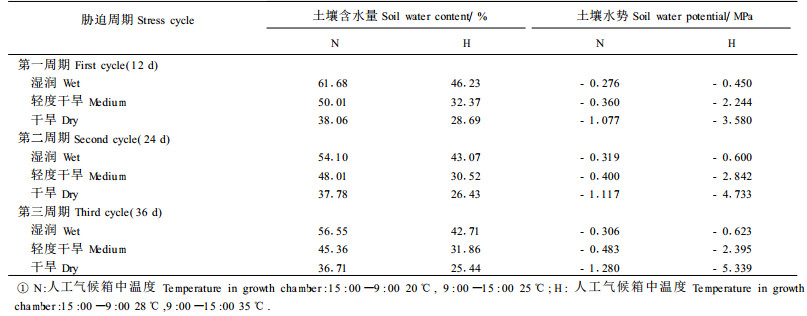
1.3 土壤水势测定在每个水分和热胁迫周期结束时,随机抽取每种水分和热胁迫处理的基质样品3份,每个周期共取土壤样品18份,总计54份,用于测定土壤水势。测定时,先选取不同干湿程度的土壤样品20份,用热电偶湿度计测定电流读数,同时测定已知水势的不同浓度NaCl标准溶液的电流读数,推导出20份被测土壤样品的水势。水势测定结束后,用105℃恒温法测定含水量,根据两者的关系,建立土壤含水量与土壤水势之间的回归方程。然后,用105℃恒温法测定每份样品的土壤含水量,利用所建立的回归方程,计算样品的水势。
1.4 苗木气体交换参数的测定在3个周期的水分和热胁迫处理结束后,用2d时间,用LCA-4型二氧化碳叶室分析系统(英格兰ADC公司)测定苗木净光合速率(Pn)和针叶气孔导度(gwv)。气体交换参数的测定在室内进行,环境气温基本保持恒定,为20℃。用支架将人工光源和光合叶室固定,保持光源与叶室间的距离恒定。测定时到达叶室的光量子通量密度为750μmol·m-2s-1。测定结束后,用叶面积仪(美国Li-COR公司)测定叶室中针叶的叶面积。根据叶面积计算修正的Pn和gwv值。计算苗木瞬间水分利用效率(Pn/gwv)。
1.5 苗木针叶碳同位素组成的测定将冷冻干燥的苗木针叶样品磨碎成粉末。每份样品称取针叶粉末1g, 用质谱仪分析样品中13C/12C,利用PDB化石的碳同位素比为标准,计算试样与标准样品偏离的千分率,即得苗木针叶碳同位素组成δ13C值。
1.6 试验设计和统计分析试验采用三因素(水分、温度和人工气候箱)随机区组设计。水分因素有3个水平,即湿润、轻度干旱和干旱。温度因素有2个水平,即28~35℃和20~25℃。人工气候箱因放置地方不同而作为一个因素作统计处理。4个人工气候箱每2个为一组,分别放置在2个地方。每种处理有8~9株苗木,选取3株苗木进行测定。采用SAS软件分析测定数据。用其中的ANOVAs程序作三因素方差分析,用DUNCAN′s程序做多重比较。
2 结果与分析 2.1 土壤水势的确定采用热电偶湿度计法,测定部分土壤样品(共20份)的水势,同时用105℃恒温法测定其含水量,据此建立土壤含水量(x)与土壤水势(y)之间的数学关系(图 1):
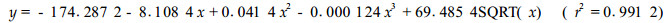
|

|
图 1 土壤含水量与土壤水势之间的关系 Fig. 1 The relationship between soil water content and soil water potential |
利用图 1建立的关系,计算样品水势(表 2)。从图 1看出,本试验所用的土壤(培养基)沙性较强,在含水量较高(50%~100%)的情况下,土壤水势降低的速度较慢。但在含水量较低(40%以下)的情况下,土壤水势下降较快。本试验采用不同浇水次数来控制苗木的水分状况,每次均将水浇透。因此,在每个胁迫周期中,浇水初期苗木水势下降较慢,后期下降较快。从表 2看出,当人工气候箱设定在28~35℃高温时,土壤水势下降幅度很大,而在20~25℃下,土壤水势下降速度较缓。说明高温会大大加速土壤失水速度。
|
|
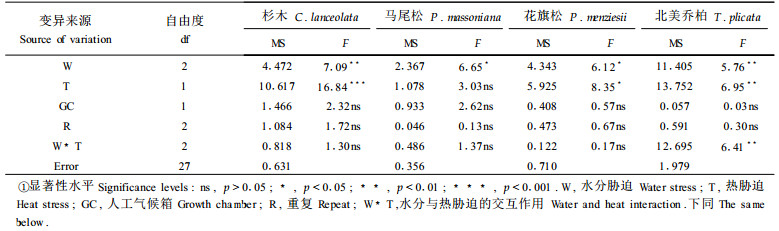
气体交换参数主要包括气孔导度、蒸腾速率、光合速率和呼吸速率等。气体交换参数是反应苗木光合作用状况的最重要指标,苗木遭受环境胁迫,其光合作用必然要受到影响,最终影响苗木的生长。为了研究水分和热胁迫对苗木气体交换参数的影响,本试验对测定条件实行了严格控制,并在36d水分和热胁迫处理结束后,立即对杉木、马尾松、花旗松和北美乔柏的净光合速率和针叶气孔导度进行了测定。结果发现,水分胁迫对4个树种的净光合速率均有显著影响(表 3),随着水分胁迫程度的加深,苗木的净光合速率下降。从净光合速率下降的幅度来看,北美乔柏最大(图 2A)。研究结果还显示,热胁迫处理只对杉木、花旗松和北美乔柏的净光合速率有显著影响,对马尾松的净光合速率影响不显著(表 3)。从影响程度来看,以杉木为最大(图 2B)。
|
|

|
图 2 36d水分和热胁迫处理对净光合速率的影响 Fig. 2 The net photosynthetic rate in response to 36 days water and heat stress treatments w:每2 d浇水一次Watering once in two days;m:每4 d浇水一次Watering once in four days;d:每6 d浇水一次Watering once in six days. n:正常温度Normal temperature:15:00—9:00 20℃, 9:00—15:00 25℃; h:高温High temperature:15:00—9:00 28℃, 9:00—15:00 35℃.下同The same below. |
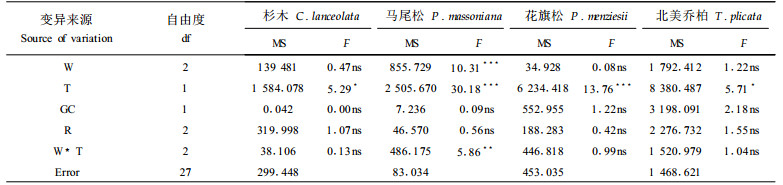
苗木针叶气孔导度是指示苗木遭受水分和热胁迫的敏感因子,也是苗木自我调节的一种生理机制。对水分和热胁迫下杉木、马尾松、花旗松和北美乔柏4个树种苗木针叶气孔导度的方差分析结果表明, 水分胁迫对马尾松苗木针叶气孔导度有显著影响,对其它3个树种的苗木针叶气孔导度影响不显著(表 4)。热胁迫则对4个树种苗木针叶的气孔导度均有显著影响(表 4),说明苗木针叶气孔导度对热胁迫的敏感程度相对高于对水分胁迫的敏感程度。从影响程度来看,水分胁迫和热胁迫均是对北美乔柏针叶气孔导度的影响程度最大(图 3),说明在4个树种中,北美乔柏苗木的针叶气孔导度对水分和热胁迫最敏感。
|
|

|
图 3 36d水分和热胁迫处理对气孔导度的影响 Fig. 3 The needle conductance to water vapor in response to 36 days water and heat stress treatments |
水分利用效率的变化是植物抗旱策略的重要组成部分。当苗木遭受水分胁迫时,其水分利用效率必定会发生相应的变化,以保持苗木生长与水分消耗的平衡。在干旱情况下,苗木的水分利用效率通常会上升,这是因为水分亏缺会导致气孔导度大大降低,从而使苗木蒸腾强度的降低大于光合强度的降低。如对黑云杉(Picea mariana)和白云杉(P.glauca)的研究表明,苗木水分利用效率与干物质产量成正比(Flanagan et al., 1995; Livingston et al., 1999)。水分利用效率依情况不同有2种表达方式:即瞬间水分利用效率和永久水分利用效率。瞬间水分利用效率是指消耗一定量的水分所同化CO2的量,可由净光合速率Pn和蒸腾速率E的比值(Pn/E)或由净光合速率Pn和气孔导度gwv的比值(Pn/gwv)求得。永久水分利用效率是指一段时间内环境参数对光合过程的影响, 通常通过测定植物组织中稳定碳同位素的组成而求得。
本试验通过测定杉木、马尾松、花旗松和北美乔柏4个树种苗木在36d水分和热胁迫后的净光合速率Pn和气孔导度gwv,计算出了4个树种苗木的瞬间水分利用效率。结果发现,水分和热胁迫处理对马尾松苗木的瞬间水分利用效率有显著影响,但对杉木、花旗松和北美乔柏的瞬间水分利用效率无显著影响(表 5)。
|
|
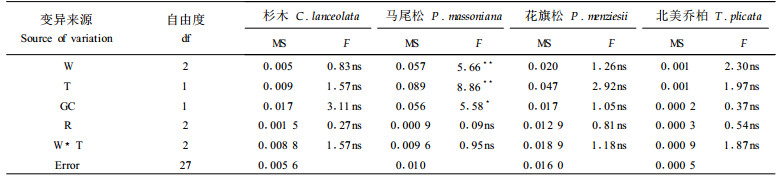
尽管方差分析显示只有马尾松苗木的水分利用效率在不同水分和热胁迫处理间存在显著差异,但图 4表明,随着水分和热胁迫加强,杉木、马尾松、花旗松和北美乔柏的瞬间水分利用效率均有下降的趋势。由于瞬间水分利用效率是由光合速率和气孔导度2个因子决定的,因此,瞬间水分利用效率的降低说明苗木在水分和热胁迫下,光合速率的下降速度大于气孔导度的下降速度。本试验的这一结论与前人结果不一致,可能因为这一测定是在瞬间进行的,难以解释苗木的长期生长情况,因此,需要测定苗木的永久水分利用效率。

|
图 4 36 d水分和热胁迫处理对瞬间水分利用效率(Pn/gwv)的影响 Fig. 4 The instantaneous water use efficiency in response to 36 days water and heat stress treatments |
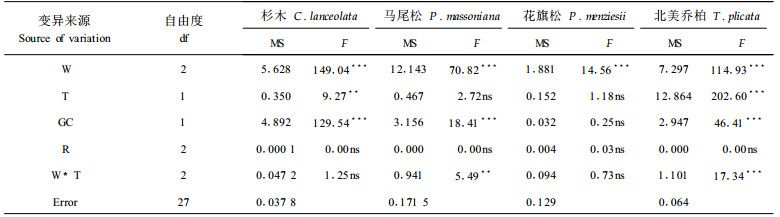
为了研究水分和热胁迫处理期间苗木永久水分利用效率状况,在36d水分和热胁迫处理结束后,对杉木、马尾松、花旗松和北美乔柏4个树种苗木针叶的稳定碳同位素组成进行了测定,结果见表 6、图 5。可以看出,水分胁迫处理对4个树种的稳定碳同位素组成(δ13C值)均有显著影响,随着水分胁迫程度的加深,4个树种苗木针叶的δ13C值增大(负值绝对值变小),这说明其永久水分利用效率更高。热胁迫对杉木和北美乔柏苗木针叶的δ13C值有显著影响,且在热胁迫下,2树种苗木针叶的δ13C值减小(负值绝对值增大),即永久水分利用效率下降。但热胁迫对马尾松和花旗松苗木的δ13C值没有显著影响。
|
|

|
图 5 36 d水分和热胁迫处理对δ13C值的影响 Fig. 5 The carbon isotope composition (δ13C) in response to 36 days water and heat stress treatments |
气体交换参数是反应苗木光合作用状况的最重要指标,苗木遭受环境胁迫,其光合作用必然要受到影响,最终影响苗木的生长。Grossnickle(2000)指出,云杉苗木遭受水分胁迫,其气体交换参数将会下降。当水势下降到低于-2.0 Mpa时,气孔导度将降为0。现已观察到,当白云杉(Grossnickle et al., 1987; 1989)、北美山地云杉(Picea engelmannii)(Lopushinsky et al., 1974; Kaufmann, 1979; Grossnickle et al., 1985)苗木在田间栽植时,由于水势的下降,其气孔会迅速关闭,净光合速率也会随水势的下降而降低。对许多云杉树种如黑云杉(P.mariana)(Patterson et al., 1997)、内陆云杉(Picea glauca×P.engelmannii)(Grossnickle et al., 1994; Fan et al., 1997)、北美云杉(Picea sitchensis)(Fan et al., 1997)和白云杉(Patterson et al., 1997)的研究也表明,当水势下降到-2.0~3.0mpa时,净光合速率降到光补偿点。本研究得到的结果与上述研究基本一致,即在水分胁迫下,4个被测树种苗木的净光合速率均显著降低。但本研究发现,不同树种的净光合速率对水分胁迫的反应不同,其中以北美乔柏对水分胁迫最为敏感,说明北美乔柏的耐旱能力最差。
高温胁迫同样会对苗木的气体交换参数产生影响。如黑云杉在23℃时,净光合速率就会显著下降;而当气温达到或大于35~40℃时,光合速率将会很低甚至停止(Meng et al., 1993; Lamhamedi et al., 1994; Vann et al., 1994; Alexander et al., 1995)。本研究的结果也证实了这一点。但高温胁迫的形式有多种,如短期的极端高温、持续的夏季高温等。本文研究的是杉木、马尾松、花旗松和北美乔柏苗木对持续夏季高温的生理反应,从结果来看,马尾松最能耐夏季持续高温,温带树种花旗松和北美乔柏则明显不耐夏季高温。
Schulze(1986)指出,叶气孔导度比光合作用对土壤水分的变化更敏感。如有些实验发现,对一些树木(Zhang et al., 1994)和作物,在水分胁迫下气孔导度下降的幅度大大高于净光合速率。在这种情况下,瞬间水分利用效率(Pn/gwv)将提高。但本实验的结果有些不同,发现叶气孔导度并不比光合作用对水分胁迫更敏感。而且还发现,在36d水分和热胁迫处理后,苗木的Pn/gwv值有轻微的下降,其中的原因值得进一步探讨。但是,这一实验结果却与Olivas-Garcia等(2000)的结果一致,这说明供试树种均有较强的调节光合作用以适应环境变化的能力,当然,这种能力是因树种而异的。本研究还发现,水分和热胁迫处理对马尾松苗木的Pn/gwv值有显著影响,可能是水分和热胁迫处理对马尾松苗木的气孔导度产生了显著影响。
Alexander J D, Donnelly J R, Shane J B. 1995. Photosynthetic and transpirational responses of red spruce understory trees to light and temperature. Tree Physiol, 15: 393-398. DOI:10.1093/treephys/15.6.393 |
Fan S, Grossnickle S, Sutton B C S. 1997. Relationship between gas exchange adaptations of Sitka×Interior spruce genotypes and ribosomal DNA markers. Tree physiol, 17: 115-123. DOI:10.1093/treephys/17.2.115 |
Flanagan L B, Johnsen K H. 1995. Genetic variation in carbon isotope discrimination and its relationship to growth under field conditions in full-sib families of Picea mariana. Can J For Res, 25: 39-47. DOI:10.1139/x95-005 |
Grossnickle S C, Reid C P P. 1985. Environment and physiological control of stomatal response of Picea engelmannii seedlings on a high elevation mine site. Ecol Plant, 6: 111-123. |
Grossnickle S C, Blake T J. 1987. Water relations and morphological development of bare-root jack pine and white spruce seedlings: seedlings establishment on a boreal cut over site. For Ecol Manage, 18: 299-318. DOI:10.1016/0378-1127(87)90133-2 |
Grossnickle S C, Heikurinen J. 1989. Site preparation: water relations and growth of outplanted jack pine and white spruce. New For, 3: 99-123. DOI:10.1007/BF00021576 |
Grossnickle S C, Major J E. 1994. Interior spruce seedlings compared to emblings produced from somatic embryogenesis. Ⅱ. Stock quality assessment prior to field planting. Can J For Res, 24: 1385-1396. DOI:10.1139/x94-179 |
Grossnickle S C. 2000. Ecophysiology of northern spruce species: the performance of planted seedlings. Ottawa: NRC Research Press.
|
Kaufmann M R. 1979. Stomatal control and the development of water deficit in Engelmann spruce seedlings during drought. Can J For Res, 9: 297-304. DOI:10.1139/x79-051 |
Lamhamedi M S, Bernier P Y. 1994. Ecophysiology and field performance of black spruce (Picea mariana): a review. Ann For Sci (Paris), 51: 529-551. |
Livingston N J, Guy R D, Sun Z J, et al. 1999. The effects of nitrogen stress on the stable carbon isotope composition, productivity and water use efficiency of white spruce (Picea glauca(Moench)Voss) seedlings. Plant Cell Environ, 22: 281-289. DOI:10.1046/j.1365-3040.1999.00400.x |
Lopushinsky W, Klock G O. 1974. Transpiration of conifer seedlings in relation to soil water potential. For Sci, 20: 181-186. |
Meng F, Arp P A. 1993. Net photosynthesis and stomatal conductance of red spruce twigs before and after detachment. Can J For Res, 32: 716-721. |
Olivas-Garcia J M, Cregg B M, Thomas C H. 2000. Genotypic variation in carbon isotope discrimination and gas exchange of ponderosa pine seedlings under two levels of water stress. Can J For Res, 30: 1581-1590. DOI:10.1139/x00-080 |
Osório J, Pereira J S. 1994. Genotypic difference in water use efficiency and 13C discrimination in Eucalyptus globulus. Tree Physiol, 14: 871-882. DOI:10.1093/treephys/14.7-8-9.871 |
Patterson T B, Guy R D, Dang Q L. 1997. Whole-plant nitrogen and water relations traits, and their associated trade-off, in adjacent muskeg and upland boreal spruce species. Oecologia, 110: 160-168. DOI:10.1007/s004420050145 |
Schulze E D. 1986. Carbon dioxide and water vapor exchange in response to drought in the atmosphere and in the soil. Annu Rev Plant Physiol, 37: 247-274. DOI:10.1146/annurev.pp.37.060186.001335 |
Sinclair T R, Ludlow M M. 1985. Who taught plants thermodynamics? The unfulfilled potential of plant water potential. Aust J Plant Physiol, 12: 213-217. |
Vann D R, Johnson A H, Casper B B. 1994. Effects of elevated temperatures on carbon dioxide exchange in Picea rubens. Tree Physiol, 14: 1339-1349. DOI:10.1093/treephys/14.12.1339 |
Zhang J W, Marshall J D. 1994. Population differences in water-use efficiency of well-watered and water-stressed western larch seedlings. Can J For Res, 24: 92-99. DOI:10.1139/x94-014 |