 2004, Vol. 40
2004, Vol. 40文章信息
- 郭志华, 肖文发, 蒋有绪.
- Guo Zhihua, Xiao Wenfa, Jiang Youxu.
- 海南岛植被景观的斑块特征
- Patch Characteristics of the Vegetation Landscape in Hainan Island
- 林业科学, 2004, 40(2): 9-15.
- Scientia Silvae Sinicae, 2004, 40(2): 9-15.
-
文章历史
- 收稿日期:2002-12-10
-
作者相关文章


植被的斑块特征是植物群落最重要的空间特征之一,斑块特征是景观空间结构的一个主要参数(肖笃宁,1991)。斑块大小与生态系统的能量流动、物质循环、物种及其他生态学现象与过程等息息相关, 斑块大小、形状和分布对动物的迁移、生物多样性的分布有重要影响(Bolger et al., 1991;Van Apeldoorn et al., 1994;McIntyre, 1995;Harrison,1997;邬建国,2000)。斑块大小和隔离度还影响物种灭绝和定居,生境的破碎化限制了小斑块中物种数量的增加(Wahlberg et al., 1996)。因此,正确理解森林斑块的大小、形状和位置(即格局)与生态学过程的关系,是合理设计森林经营策略的基础(Pan et al., 2001)。另外,岛屿生物地理学认为:在距离相同的条件下,岛屿面积越大,岛上的物种数量越多(Wardle et al., 1997),而缓冲区的面积太小也是保护区内大型肉食动物消失的重要原因(Woodroffe et al., 1998)。在自然保护区规划设计时,保护区的大小是最主要的考虑因素。所以,研究景观斑块的大小特征有利于保护区的规划设计。
过去描述斑块形状的指标通常是形状指数,但形状指数不能提供斑块形状复杂性的直接度量(刘灿然,2000)。近年来,常用分维方法来进行生态学的格局分析(Mladenoff et al., 1993;Cain et al., 1997; 王宪礼等,1997;Nikora et al., 1999;刘灿然,2000)。利用专题地图来研究景观斑块大小特征及其分形是常用的手段(郭晋平等,2000b; 马克明等,2000;刘灿然,2000)。已收集的海南岛相关专题地图有:1:10万土地利用图,1:20万林相图和1:50万植被图。由于海南岛土地利用图仅仅将森林景观划分为林地和非林地, 而林相图仅根据林相分类, 所以,这两种数据均不能满足植被景观的研究所需。于是,本研究选择由中国科学院华南植物研究所绘制的海南岛植被图作为植被景观斑块研究的数据源。
中国科学院华南植物研究所(1989)在1980—1988年间经地面样地调查,借助于卫片、航片等绘制了广东省植被类型图(包含海南岛植被)。海南岛植被类型包括热性针叶林、阔叶林、灌丛、草丛、农业植被、人工林和水生植被等7大类型。其中阔叶林又包括常绿阔叶林、季雨林、雨林、红树林、竹林等5个类型。上述7大类型植被景观分别包含1、12、7、5、4、6和1个基本类型, 共计36个基本类型植被景观。利用ARC/INFO和数字化仪将植被图数字化以提取斑块信息。相临的基本类型斑块通过去除公共边而生成7大类型植被斑块。
研究海南岛植被斑块的大小及分形特征,评价不同类型、不同尺度上的植被斑块复杂性,既可为探讨海南岛景观格局的动态变化规律奠定基础,也可为海南岛景观水平上的生物多样性保护提供理论基础。
1 研究方法 1.1 斑块大小特征的研究方法植被景观斑块的大小特征及分布规律采用以下统计量来描述:斑块数、平均斑块面积、斑块面积标准差、斑块总面积、最小斑块面积、最大斑块面积、斑块面积极差、斑块面积分布的偏度系数、斑块面积分布的峰度系数、斑块面积的中位数和众数等。
本文选择了正态分布、Gamma分布、对数正态分布、Weibull分布、指数分布等来检验海南岛植被斑块大小分布的概率分布类型。离散数据的分布相似性检验用Smirnov方法。
1.2 斑块分形的研究方法单个斑块的分维数从下式求得(Frohn, 1998):
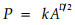
|
(1) |
式中,P是斑块的周长,k是常数,A是斑块的面积,D为分维数。
对于同一类斑块的分维数,均采用最小二乘法,通过求回归系数的方法获得。
方法1:
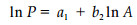
|
(2) |
方法2(李哈滨,1992; Klinkenberg, 1994):
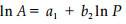
|
(3) |
(2) 式中,a1为该直线在lnP轴上的截距, b1为该直线的斜率(Krummel et al., 1987),分维数D1=2b1;(3)式中,a2为直线在lnA轴上的截距,b2为该直线的斜率,分维数D2=2/b2。利用(2)和(3)式分别计算了海南岛不同等级类型植被斑块的分维数,并分析了分维数与斑块大小特征之间的相关性,检验了不同等级类型植被斑块分维数的差异显著性,进而探讨了海南岛植被斑块的尺度域问题。所有数量统计分析均在SAS软件下进行。
2 结果 2.1 斑块大小36个基本类型的统计量计算结果表明:水稻群落(Comm. Oryza sativa)的斑块数最多(65), 占总斑块数的12%以上;最少斑块数为1, 如海南锥-竹叶木姜林(Comm.Castanopsis hainanensis+Litsea pseudoelongata)、银叶树林(Comm. Heritiera littoralis)等。平均面积最大的是青皮-山荔枝林(Comm. Vatica astrotricha+Litchi chinensis var. euspontanea)(达247.0km2),最小的是银叶树林(仅0.5km2),两者相差约500倍。标准差最大的是青皮-山荔枝林。总面积最大的是水稻群落,达5 287.9 km2;最小的是银叶树林。在各基本植被类型中,最小面积(MN)最大的是桃金娘灌丛(Comm.Rhodomyrtus tomentosa)(达112.3km2),最小的是红海榄林(Comm.Rhizophora stylosa)和银叶树林(为0.5km2);最大面积(MX)最大的是水稻(为1 384.3 km2);变异系数(CV)以水稻群落最大(为238.4);面积极差(RG)最大的也是水稻群落(达1 382.0km2);从偏度系数(SK)来看,除斑块数少(<4)的类型外, 多数类型植被斑块的偏度系数>0,仅鸡毛松林(Comm. Podocarpus imbricatus)、青皮竹林(Comm. Bambusa textilis)为负。这表明多数植被类型斑块大小的分布是正偏的,并且绝大多数斑块面积的中位数MD和众数MO都小于其平均面积。从峰度值(KT)来看,大多数类型斑块的KT值为正,少数为负,表明这些类型斑块大小的分布曲线是尖峭峰,且以水稻群落的KT值为最大(33.7)。
在阔叶林斑块中,海南岛季雨林的斑块数最多,其平均斑块面积、标准差、总面积、最大斑块面积、斑块面积极差、偏度系数、中位数等最大。这说明,季雨林在海南岛的分布很广且破碎。雨林的最小斑块面积、变异系数和众数等值最大。红树林的峰度系数最大。除竹林外,其他阔叶林斑块面积的分布曲线都是正偏态;常绿阔叶林和竹林斑块面积的分布曲线为平阔峰,其余为尖峭峰。由于红树林仅仅分布在淤泥质的海滩,因此,其平均斑块面积最小。竹林的平均斑块面积也不大(10.0km2), 这些都与人类活动的影响有关。
从7大类型植被斑块的上述统计量(表 1)可以看出,灌丛和农田的斑块数量最多,水生植被的斑块数量最少。农田斑块的平均面积和总面积均最大,这表明海南岛的土地利用方式以农耕为主,且多集中成片。此外,农田植被的最大斑块面积、斑块面积的变异系数、极差、中位数、偏度值和峰度值在各大类型中均最大。农田的总面积为10 153km2,约占海南岛面积的30%。其次是灌丛(7 090.9km2)、阔叶林(6 095.3km2)、人工林(6 065.8km2)和草丛(4 331.9km2),大约分别占海南岛面积的21%、18%、18%、18%和13%。热性针叶林和水生植被的面积很小,均不及海南岛面积的0.5%。虽然海南岛的森林约占岛屿的36%,但天然林(阔叶林)所占比重不大,且阔叶林斑块大小的变异很大,其SD=119.0,CV=177.7,最大斑块面积是最小斑块面积的1 400倍左右,这除了与地形破碎有关外,还表明了海南岛森林植被深受人类活动影响。海南岛少数民族多有刀耕火种的农作习惯,致使大片森林退化为灌丛和草丛,严重地影响了当地的林业发展。各大类型斑块大小的偏度值和峰度值均为正,表明这些斑块面积的分布都是不对称的,并且都是正偏(即右偏)和尖峭峰,其中农田斑块面积的分布曲线最右偏和最陡峭。
|
|
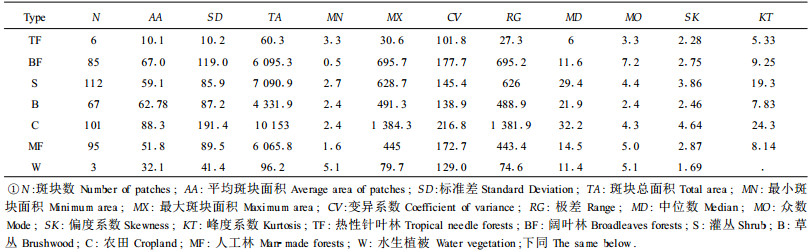
为了更好地反映各大类型斑块大小的分布特征,将斑块大小分为31个等级,它们分别是<5、[5, 10)、[10, 20)、[20, 30)、[30, 40)……[280, 290)、≥290, 单位是km2(图 1)。由于热性针叶林和水生植被的斑块数量少,因此仅讨论其余5大类型斑块的大小分布特征。从图中可知,5大类型植被景观均以<10km2的斑块数量最多。阔叶林在第一个等级(<5km2)上的斑块数最多,灌丛、草丛和农田在第三等级[10~20km2)上的数量最多,而人工林却在[5~10km2)的数量最多。大斑块的数量虽少,却占有很大比例的面积,尤其以阔叶林、农田和人工林等类型更明显。从斑块大小的分布来看,各大植被类型的分布显然都是正偏的(图 1)。相对地,草丛斑块面积的变化较小,人工林斑块面积的变化最大。对数量大于30的5大类型植被斑块进行分布函数的拟合检验,结果见表 2。从表 2中可知,这5大类型植被斑块的分布都不是正态分布和指数分布,并且本文所选的分布函数均不能反应人工林斑块大小的分布特征。相对地,对数正态分布能较好地反应各大类植被斑块的分布特征。阔叶林斑块大小的分布对Gamma和Weibull分布的拟合较好。

|
图 1 5大类型斑块的频数分布和面积分布(面积单位为km2) Fig. 1 Patch frequency distribution and patch area distribution for the 5 higher-level types(area unit is km2) |
|
|

对5大类型植被斑块分布函数的相似性进行Simirnov检验(表 3),结果表明:仅阔叶林和人工林、灌丛和农田之间无显著差异外,其余类型之间均存在显著或极显著差异。阔叶林与人工林之间无显著差异显示了森林植被的共同属性,而灌丛与农田斑块大小分布的相似性说明两者受到人类活动深刻影响。
|
|

用两种方法计算海南岛36个基本植被类型的分维数。结果表明:当斑块数较少时,均出现分维数小于1或大于2的异常情况,效果欠佳;用方法2计算的分维数比用方法1计算的分维数稍大,但当斑块数较多时,计算结果的差异不大,因此,在斑块数较多时用这两种方法均可获得对斑块分维数的可靠估计。
对于数量大于20的11个基本类型,其分维数D1与D2之间具有很强的正相关性(r=0.960 7, p<0.01)。斑块的分维数D1及D2与其描述性统计量之间的相关性见表 4。从表 4可知,分维数D1与各项统计量之间均没有显著的相关性(p<0.1),但D2却与AA、SD、MX、RG和SK等之间存在不同程度的相关性。分维数D2随斑块平均面积的增加而增加(0.05<p<0.1),随斑块面积标准差的增加而降低(p<0.01),随最大斑块面积的增加而减小(0.01<p<0.05),随斑块面积极差的增大而降低(0.01<p<0.05),随偏度值的增加而降低(0.05<p<0.1)。对斑块数大于20的11个基本类型植被斑块的分维数进行两两之间的差异显著性检验。结果表明,山地雨林、丘陵山地草丛、水稻群落和木麻黄林等植被斑块的分维数与多数类型植被斑块分维数之间有显著差异,而其余类型大多与别的斑块分维数之间无显著差异。相反,并未发现同一大类型植被斑块之间分维数差异不显著的现象。
|
|
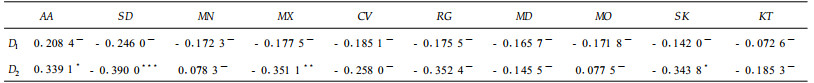
7大类型植被斑块的分维数及相关统计量见表 5。从表中可知,lnA和lnP之间的相关系数都很大,都达到极显著水平(p<0.01);水生植被的分维数小于1,这是因为其斑块数很少的缘故;热性针叶林的分维数最大,其余依次是阔叶林、草丛、灌丛和人工林,农田的分维数很小。这种分维数的变化特征与北京植被斑块分维数的变化特征略有不同(刘灿然等,2000)。
|
|

对数量大于20的5大类型植被斑块的分维数两两之间进行差异性检验,结果表明,除灌丛与阔叶林、灌丛与人工林及草丛与农田之间,它们形状的复杂性无显著差异外,其余两两之间均有显著或极显著差异。
2.3.3 尺度域的确定从5大类型植被斑块lnA与lnP的散点图(图 2)可知,在lnA的定义域内,各大类型植被斑块lnA和lnP之间不是简单的线性关系,特别是对阔叶林斑块而言。这就表明,海南岛各大类型植被斑块在不同尺度域内的分维数不同,其转折点大约在lnA=2附近(图 2)。

|
图 2 5大类型群落斑块周长(P)和面积之间的关系(A) Fig. 2 Relationship between perimeter(P) and area (A) for the patches of 5 higher-level types ——、——分别用lnA>2、lnA<2的点拟合的直线 ——、——Line fitted to the data with lnA>2 and A<2 respectively.D2和D'2分别代表用方法2计算的lnA>2及lnA<2的群落斑块的分维值D2, D′2:estimated fractal dimension for the patches with lnA>2 and lnA<2 respectively based on the method 2. |
将5大类型植被的所有斑块,按lnA<2和lnA>2分成两部分,应用(2)式,再分别求各大类型植被每部分的分维数,并进行差异显著性检验。结果表明,各大类型的大、小斑块的分维数之间均有显著差异。因此,可以将5大类型植被斑块按lnA=2将尺度域分成两部分,这表明:在斑块面积A=e2≈7.4(km2)左右时,斑块形状的复杂性发生了显著变化(p<0.05)。
3 讨论 3.1 关于斑块的分类、界定景观生态学研究的一个基本出发点就是斑块的界定,即斑块的分类和划界。但是,关于斑块的定义迄今争议不休。通常地,斑块就是“在性质或表面上与周围明显不同的区域”(福尔曼等,1990)。而很多学者却认为,斑块可以根据研究的内容和要回答的问题来定义(Kotliar et al., 1990)。景观生态学是在20世纪60年代从地理学和生物地理学中分支出来的(福尔曼等,1990)。从诞生的那天起,景观生态学似乎就成了地理学家和生物地理学家的专利,他们真正关注的是土地利用方式而不是在地球陆地表面占绝对优势的植物群落。
在一定程度上,不同的景观斑块就是不同的植被类型,然而植被分类也充满争议。不同学派根据不同的分类原则提出了不同的分类系统。中国植被的分类有其独立的分类原则和分类系统,而海南岛的植被分类又与中国植被的分类系统有所不同。所以,如何科学地对陆地植被进行准确分类、制图是大尺度景观生态学研究的关键。由于海南岛的农业用地很多,因此,本文充分重视了人类活动对海南岛植被分类的影响。
但是景观斑块又不等同于植物群落,因为景观生态学很重视廊道和基质的作用。道路、河流渠等廊道对景观破碎化的影响很大,特别是在平原、城市等人类活动密度大的地区。然而在植物群落的分类制图时几乎不考虑这些因素的影响。因此,仅仅利用植被图在宏观尺度上研究景观斑块特征还存在一定局限。今后的工作若能叠加道路、河流等的影响,则或许会取得更好的效果。如果这样,农田斑块就不再有巨大面积,其大小的分布特征及分形也将有所不同。在大尺度上进行景观分类、景观格局分析及其他研究,遥感技术是一有效手段。近年来,遥感技术和方法在景观生态学研究中的应用越来越广,并取得了很好的研究效果(Debinski et al., 1999;Chuvieco,1999;彭少麟等,1999;郭晋平等,2000a;2000b;曾辉等,2000;吴波等,2001;Pan et al., 2001),并将继续在景观生态学研究中居重要地位。
3.2 关于地图精度与研究尺度植被景观图可以分为矢量和栅格两大类型。传统地图是矢量数据,现在的大多数景观斑块特征和景观格局研究多基于那些传统的专题地图。这样,研究精度就取决于地图精度。不同比例尺地图表达地物的精度不同,因此,选择适当比例尺的地图是很重要的,这取决于研究尺度的大小。当然,除了考虑比例尺大小外,还应考虑成图质量和研究所需。当景观斑块很小时,图斑边界的细小误差就会引起斑块分维数的较大波动。对于传统地图来讲,这种误差客观存在。此外,在传统地图制图时,还可以根据研究需要,适当强调某些在空间上很小的地物,并将其适当放大以示重要,然而这种很小的夸张将会影响景观生态的研究结果。如红树林是一类重要的植被类型,仅见于海南岛东部淤泥质海岸、分布面积狭小。在海南岛,在有红树林的地区就在地图上加以标识,而岛内许多比这些小斑块大得多的植被斑块未表示出来。所以,上述基于海南岛植被图所做研究而得出的大斑块的形状比小斑块的形状复杂的结果还有待进一步验证。
近年来,利用遥感数据所进行的景观生态学研究已越来越多,这样的景观斑块特征和格局分析研究都是基于栅格数据的,因此,研究尺度问题与栅格数据的分辨率直接相关。然而,自然景观本身不可能是栅格的。所以,在利用遥感数据进行景观生态学研究时,必须重视其空间分辨率的大小对研究结果的影响,并且根据研究尺度和研究需要来选择不同分辨率的遥感数据。
景观生态学认为,具有分形结构的景观,其斑块在不同尺度上应该表现出很大的相似性。通过在不同尺度域上计算其分维数,观察其值的变化,可以显示和判断景观的等级结构。如果分维数在某一尺度范围内保持不变,则景观在这一尺度范围内具有结构相似性;如果分维数在不同的尺度域发生了变化,则发生变化的转折点就指示了景观的等级结构(Leduce et al., 1994)。本文的初步研究结果表明,面积<7.4km2和面积>7.4km2的斑块具有显著不同的分维数(p<0.05),这是基于1:50万的植被图得出的。当然,若利用不同比例尺的地图或利用TM数据对海南岛景观分类,则海南岛植被景观斑块的尺度域可能不同。
4 小结海南岛基本类型植被景观的斑块数介于1~65之间,面积介于0.5~1 384.3km2之间。
阔叶林约占海南岛景观面积的18%。其中,季雨林分布最广,其次是雨林,其余类型的面积很少。
从各大类型植被斑块来看,灌丛和农田的斑块数量最多;农田的总面积最大,其次是灌丛、阔叶林、人工林和草丛,这表明海南岛的土地利用方式还是农耕为主。在海南岛森林中,天然林所占比重不大。
对于数量较多的基本类型斑块,海南岛植被斑块的分维数D2与斑块平均面积呈正相关,与斑块面积标准差、最大斑块面积、斑块面积极差、偏度值呈负相关。
7大类型植被斑块分维数从大到小依次是热性针叶林阔叶林、草丛、灌丛、人工林、农田和水生植被。除灌丛与阔叶林、灌丛与人工林及草丛与农田之间的分维数无显著差异外,其余两两之间均有显著或极显著差异。
对于斑块数较多的5大类型植被,以斑块面积7.4km2为界,斑块形状的复杂性将发生显著变化。
陈利顶, 傅伯杰, 刘雪华. 2000. 自然保护区景观结构设计与物种保护——以卧龙自然保护区为例. 自然资源学报, 15(2): 164-169. DOI:10.3321/j.issn:1000-3037.2000.02.013 |
郭晋平, 王俊田, 李世光. 2000a. 关帝山林区景观要素沿环境梯度分布趋势的研究. 植物生态学报, 24(2): 135-140. |
郭晋平, 薛俊杰, 李志强, 等. 2000b. 森林景观恢复过程中景观要素斑块规模的动态分析. 生态学报, 20(2): 218-223. |
李哈滨, 武业钢. 景观生态学的数学方法. 见: 刘建国主编, 现代生态学进展. 北京: 科学技术出版社, 209-233
|
刘灿然, 陈灵芝. 2000. 北京地区植被景观斑块形状的分形分析. 植物生态学报, 24(2): 129-134. DOI:10.3321/j.issn:1005-264X.2000.02.001 |
马克明, 傅伯杰. 2000. 北京东灵山景观格局及破碎化评价. 植物生态学报, 24(3): 320-326. DOI:10.3321/j.issn:1005-264X.2000.03.013 |
彭少麟, 郭志华, 王伯荪. 1999. RS和GIS在植被生态学中的应用及其前景. 生态学杂志, 18(5): 52-64. DOI:10.3321/j.issn:1000-4890.1999.05.010 |
王宪礼, 肖笃宁. 1997. 辽河三角洲湿地的景观格局分析. 生态学报, 17(3): 317-324. DOI:10.3321/j.issn:1000-0933.1997.03.016 |
邬建国. 2000. 景观生态学——格局、过程、尺度与等级. 北京: 高等教育出版社, 25-30.
|
吴波, 慈龙骏. 2001. 毛乌素沙地景观格局变化研究. 生态学报, 21(2): 191-196. DOI:10.3321/j.issn:1000-0933.2001.02.003 |
肖笃宁. 景观的空间结构指数及其分析方法. 见, 景观生态学: 理论, 方法和应用(肖笃宁主编). 北京: 中国林业出版社, 1991
|
曾辉, 姜传明. 2000. 深圳市龙华地区快速城市化过程中的景观结构研究——林地的结构和异质性特征分析. 生态学报, 20(3): 376-383. |
中国科学院华南植物研究所. 1989. 广东省植被图(1: 500 000). 北京: 学术期刊出版社.
|
R福尔曼, M戈德罗恩(肖笃宁, 张启德, 赵羿等译). 景观生态学. 北京: 科学出版社, 1990: 53-56
|
Bolger D T, Alberts A C, Soule M E. 1991. Occurrence patterns of bird species in habitat fragments: samples, extinction, and nested species subsets. The American Naturalist, 137: 155-165. DOI:10.1086/285151 |
Cain D H, Riitters K, Orvis K. 1997. A multi-scale analysis of landscape statistics. Landscape Ecology, 12: 199-212. DOI:10.1023/A:1007938619068 |
Chuvieco E. 1999. Measuring changes in landscape pattern from satellite images: short-term effects of fire on spatial diversity. Int J Remote Sensing, 20(12): 2331-2346. DOI:10.1080/014311699212056 |
Debinski D M, Kindscher K, Jakubauskas M E. 1999. A remote sensing and GIS-based model of habitats and biodiversity in the Greater Yellowstone Ecosystem. Int J Remote Sensing, 20(17): 3281-3291. DOI:10.1080/014311699211336 |
Frohn R C. 1998. Remote sensing for landscape ecology: new metric indicators for monitoring, modeling, and assessment of ecosystems. Boca/ Raton/ Boston/ London/ New York/ Washington, D. C: Lewis Publishers, 1-94.
|
Harrison S. 1997. How natural habitat patchiness affects the distribution of diversity in Californian serpentine chaparral. Ecology, 78(6): 1898-1906. DOI:10.1890/0012-9658(1997)078[1898:HNHPAT]2.0.CO;2 |
Klinkenberg B. 1994. A review of methods used to determine the fractal dimension of linear features. Mathematical Geology, 26: 23-46. DOI:10.1007/BF02065874 |
Krummel J R, Cardner R H, Sugibara G, et al. 1987. Landscape patterns in a disturbed environment. Oikos, 48: 321-324. DOI:10.2307/3565520 |
Leduce A, Prairie Y T, Bergeron Y. 1994. Fractal dimension estimates of a fragmented landscape: sources of variability. Landscape Ecology, 9: 279-286. DOI:10.1007/BF00129239 |
McIntyre N E. 1995. Effects of forest patch size on avian diversity. Landscape Ecology, 10: 85-99. DOI:10.1007/BF00153826 |
Mladenoff D J, White J M A, Pastor J, et al. 1993. Comparing spatial pattern in unaltered old-growth and disturbed forest landscapes. Ecological Application, 3: 294-306. DOI:10.2307/1941832 |
Nikora V I, Pearson C P, Shandar U. 1999. Scaling properties in landscape patterns: New Zealand experience. Landscape Ecology, 14: 17-33. DOI:10.1023/A:1008077914643 |
Pan D, Domon G, Marceau D, Bouchard A. 2001. Spatial pattern of coniferous and deciduous forest patches in an Eastern North America agricultural landscape: the influence of land use and physical attributes. Landscape Ecology, 16: 99-110. DOI:10.1023/A:1011167905097 |
Van Apeldoorn R C, Celada C, Nieuwenhuizen W. 1994. Distribution and dynamics of the red squirrel (Sciurus vulgaris L.) in a landscape with fragmented habitat. Landscape Ecology, 9: 227-235. DOI:10.1007/BF00134749 |
Wahlberg N, Moilanen A, Hanski I. 1996. Predicting the Occurrence of Endangered Species in Fragmented Landscapes. Science, 273: 1536-1538. DOI:10.1126/science.273.5281.1536 |
Wardle D A, Zackrisson O, Hörnberg G, et al. 1997. The Influence of Island Area on Ecosystem Properties. Science, 277: 1296-1299. DOI:10.1126/science.277.5330.1296 |
Woodroffe R, Ginsberg J R. 1998. Edge Effects and the Extinction of Populations Inside Protected Areas. Science, 280: 2126-2128. DOI:10.1126/science.280.5372.2126 |