 2004, Vol. 40
2004, Vol. 40文章信息
- 李根前, 赵粉侠, 李秀寨, 韦宇.
- Li Genqian, Zhao Fenxia, Li Xiuzhai, Wei Yu.
- 毛乌素沙地中国沙棘种群数量动态研究
- Density and Biomass Dynamics of Hippophae rhamnoides L.subsp. sinensis Population in Mu Us Sandland
- 林业科学, 2004, 40(1): 180-184.
- Scientia Silvae Sinicae, 2004, 40(1): 180-184.
-
文章历史
- 收稿日期:2001-12-18
-
作者相关文章


2. 西南林学院 昆明 650224
2. Southwest Forestry College Kunming 650224
沙棘不仅是优良的多用途树种(multiple purpose tree),而且无性繁殖能力极强,能通过串根萌蘖而“独木成林”并赖以维持群落的持久性,在黄土高原生态环境建设和区域经济发展中起着积极的作用。我国拥有沙棘属植物资源92万km2, 中国沙棘(Hippophae rhamnoides L. subsp. sinensis)占88.8%,其重要性由此可见一斑。由于中国沙棘是强阳性树种,因此通常被描述为植物群落演替过程中的先锋种群(朱志诚,1992)。但在毛乌素沙地的研究表明,中国沙棘可以通过无性系生长调节实施林窗更新恢复其稳定性或依靠林缘使种群在更大的空间尺度上进行自我维持(李根前等,2001a;2001b;2001c)。本文是前述研究的继续,试图从种群数量(密度和生物量)动态方面,进一步揭示中国沙棘群落的稳定性机制,期望为中国沙棘营林措施的制定提供理论依据。
数量动态是植物种群生态学研究的核心内容之一,也是制定营林措施的重要依据,它主要描述生物量与密度的时空变化及其相互关系。一般情况下,随着密度的增加,植物个体以死亡、降低生长量或生殖能力作出反应(Harper,1981)。但在具有无性系生长的植物种类中,种群具有由遗传单位组成的母株种群和由无性繁殖所形成的构件种群等多层次水平,它们的数量变化有着不同的遗传后果(Harper,1981;Hartnett et al., 1985a;1985b;李根前等,2001a)。这方面现有的研究对象主要为1、2年生和多年生草本植物,对木本植物的研究极少报道(刘庆等,1995)。本文从基株种群(ramet population)、母株种群(mother ramet population)、子株种群(daughter ramets population)等层次上,探讨了中国沙棘的数量动态规律,可丰富木本无性系植物(clonal plant)种群生态学的研究内容。
1 研究区概况及研究方法 1.1 研究区概况研究区设在陕西省靖边县沙石峁林场和靖边县治沙站,位于毛乌素沙地南缘,属中温带半干旱大陆性季风气候,年平均气温7.8℃,年均降水量395.4?mm;土壤有风沙土、沙质草甸土和轻度盐化草甸土;地带性植被属干草原,其植物区系组成兼具旱生沙生、旱生草原及中生草甸的种类成份。中国沙棘常以建群种组成天然群落,主要分布于沙区丘间地、低湿滩地、河滩及河旁阶地等以干草原为背景的隐域性生境中,伴生灌木主要有沙柳(Salix psammphila)、毛乌柳(Salix microstachva)等,草本及半灌木层优势度较大者有黑沙蒿(Artemisia ordosica)、沙米(Agriophyllum squarrosum)、沙蓬(Sasola ruthenica)、沙竹(Psammchloa mogolica)、芦苇(Phragmites communis)、寸草(Carex sthenopylla)等。
1.2 研究方法(1) 基株种群与构件种群数量动态 基株(genetic individual)是同一颗种子衍生的所有分株的总称,是一个遗传单位;构件种群则包括叶片种群、枝条种群等。基株种群数量(密度和生物量)动态调查采用典型抽样,要求样地代表群落的不同演替阶段、立地条件相对一致、优势植物种类组成基本相似,幼林样地面积为5 m×5 m或10 m×5 m,成林为10 m×10 m。样地确定后,对样地内植株进行每木检尺,即逐株伐倒后查数年轮,并测定高度、地径(高度小于2.5 m、胸径(高度大于2.5 m)等指标。生物量测定采用径阶标准木和分层切割法,即根据样地每木检尺资料按胸径(或地径)和树高选择各径阶标准木,以1 m为一个区分段称取树干、枝条、叶片和果实的重量,对各个组分(构件种群)采集一定数量的样品置于85 ℃下烘至恒重,由此推算生物量干重。基株种群的年龄,即种群侵入立地以来的时间用样地中最大个体的年龄估计,种群数量动态的分析采用“样地编年序列法”(董鸣,1986)。(2)母株种群与子株种群密度动态 为了解母株(来自种子繁殖)种群和子株(来自无性繁殖)种密度动态,对处于不同群落演替阶段的样地C2、C3、C4(年龄结构分别为增长型、稳定型和始衰型)进行挖掘。其中,对样地C2所有基株(无性系)进行“跟踪挖掘”,即沿萌蘖根方向逐一挖出每个无性系分株(包括所有的基株与子株),同时挖掘所有的独立植株。通过全部挖掘找出所有的连结关系,基株中年龄最大者以及独立的植株即为母株,无性系中其余的植株计为子株。通过对样地C2所有个体的挖掘,利用年龄确定母株定居的时间以及子株的出生时间。根据挖掘结果,样地C2内无性系分株只有极少量的死亡,同时该样地基株种群具有稳定增长的年龄结构(李根前等,2001)。因此,可以参考“倒逐年龄累加法”(刘庆等,1996)“追踪”种群密度动态过程,如样地C2的种群年龄为6?a,则第1年(初始种群)由6 a个体组成,第2年的种群由6 a和5 a个体共同组成,其余依此类推。样地C3、C4各挖掘一个无性系,其母株种群与子株种群密度根据无性系大小及基株种群密度推算,如样地C3的基株种群密度为17 800株(丛)·hm2, 所挖掘的无性系有11个根蘖植株(子株)。那么,样地中应有母株1 483株(丛)·hm2、子株16 317株(丛)·hm2。
本次调查样地12块(C1-C12),具体情况见参考文献(李根前等,2001a)。
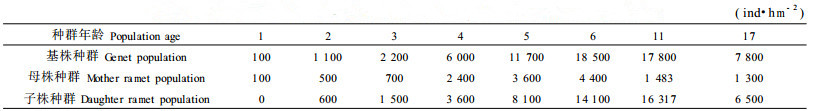
2 结果与分析 2.1 基株种群密度动态基株种群密度(D)与其年龄(A)之间的关系分两个阶段描述,5 a以前的动态依据样地C2“跟踪挖掘”资料以“倒逐年龄累加法”的统计结果(表 1)进行分析;5 a以后的动态是在12块样地中,选择6个属于不同群落演替阶段或同一阶段但种群年龄不同的样地(C1~C6)进行分析。用幂函数对样地的A、D进行拟合,得到图 1及回归方程:
|
|

|
图 1 中国沙棘种群密度动态 Fig. 1 The density dynamics of Hippophae rhamnoides L. subsp. sinensis population |
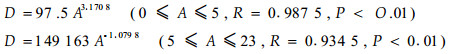
|
从回归方程和图 1可以看出,中国沙棘侵入裸地后,基株种群密度在5 a以前随年龄增大而迅速增长,6~10 a期间随年龄增大而急剧下降。此后,种群密度缓慢下降(10~20 a),然后保持在一定水平上。随着林窗的出现,种群密度又出现回升现象(C7)。
中国沙棘以种子进行迁移,当种子侵入裸地后,依靠种子萌发和极强的根蘖繁殖迅速形成含沙棘幼苗幼树的草灌丛。由于幼苗幼树个体小,立地能容纳的个体数量多,种群密度逐年增大,约在5 a时达到最大值(图 1)。随着个体的生长,林冠逐渐郁闭,种内竞争导致的种群自疏作用产生,死亡率增加,使种群密度急剧下降(6~10 a)。此时还有少量的其它一些阔叶树种(如沙柳、小叶杨)侵入,它们与沙棘发生种间竞争引起他疏,其结果仍可导致沙棘种群密度的下降,但这种作用自始至终是比较微弱的。随着中国沙棘种群密度的下降,通过种内和种间竞争及调节,自疏和他疏作用有所减弱,种群密度的下降缓慢下来(11 ~20 a),最后趋于保持在一定水平上。此时群落中的一部分个体处于林冠亚层或下层,由于林下光照条件极差,使得这部分个体很难生长到达林冠层,死亡率极高,但由于种群可以不断得到来自根蘖所产生的幼苗补充,所以种群密度基本仍可维持稳定。一旦大树死亡或倒伏,林下现存的中国沙棘个体能够迅速生长并补充到林冠层中去,以维持中国沙棘在群落中的优势地位。同时,林窗的出现改善了局部地段的光照条件,可以促进根蘖苗的产生,从而使种群密度回升(图 1中样地C7)。通过林窗效应,中国沙棘种群即可进行种群结构的自我修复,修复后种群密度又会下降,这种密度的波动性保证了中国沙棘在群落中的稳定性。
2.2 母株种群与子株种群密度动态无性系植物种群密度动态是母株种群和子株种群数量消长的结果,他们的数量变化具有不同的遗传后果,母株的出生与死亡进程影响母株种群大小和基因组成,而子株的出生与死亡影响母株的大小和适合度。为了解中国沙棘母株种群与子株种群的密度动态,对处于不同演替阶段的样地C2、C3、C4进行数据整理,结果见表 1。
由表 1可以看出,中国沙棘基株种群的密度变化有两个过程在起着作用,第一是母株的出生和死亡,第二是存活母株的无性系生长(根蘖营养繁殖)调节,即子株的出生与死亡(李根前等,2001a)。在6 a以前,母株的出生率大于死亡率,母株种群密度随种群年龄的增大而增长;6 a以后,母株的死亡率大于出生率,母株种群密度随年龄的增长而降低。对于子株种群而言,在11 a以前,子株的出生率大于死亡率,子株种群密度随种群年龄的增长而增大;11 a以后,子株的死亡率大于出生率,子株种群密度随年龄的增长而减小。由此表明,在中国沙棘基株种群密度变化过程中,不仅在基株之间发生了自然稀疏,而且在基株内部的分株之间也发生了稀疏。同时表明,子株种群的密度变化与母株种群相比具有滞后效应,即当母株种群密度已经开始下降时,子株种群的密度在一段时间内仍继续保持上升。而且,在基株种群密度变化过程中,母株种群数量所占的比例越来越小,而子株种群数量所占的比例越来越大。这种动态特征必然会导致中国沙棘种群的组成由较多拥有较少子株的无性系向较少拥有较多子株的无性系转变,从而降低了中国沙棘种群的基株多样性。
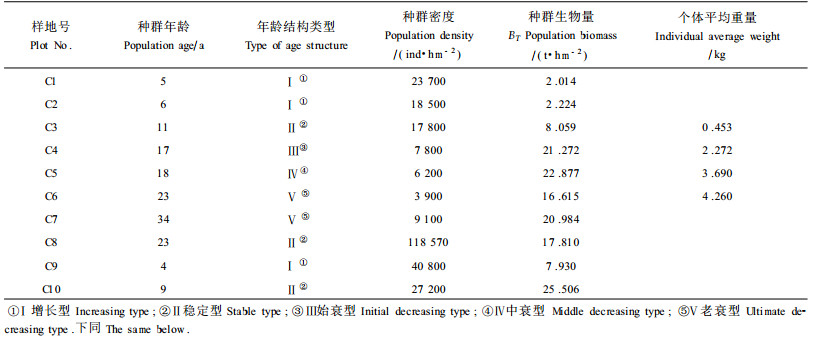
2.3 种群生物量动态现将10块样地中国沙棘种群(此处指基株种群)的地上生物量列入表 2,分析种群生物量随种群年龄的变化过程。从理论上讲,植物种群生物量积累符合Logistic增长,其表达式为:BT=BK(1+Ce-rA)-1。式中,BT为种群生物量(t·hm2);BK是立地可负荷的最大种群生物量,单位同BT;A是种群年龄(a);C和r是待定系数。参考表 2中国沙棘种群生物量变化过程,估计BK为25 t·hm2。应用样地C1~C5的资料,根据最小二乘法原理,拟合得回归方程:BT=25 (1+90.712 9e+-0.371 1A)-1(R=0.995 1,P<0.01),其图像见图 2。图 2表明,从种群增长(C1)到中衰(C5)阶段,中国沙棘种群的生物量积累符合Logistic增长,C6、C7预示了老衰阶段的生物量波动情况。根据对上述回归方程分析(李秋元等,1993),结合图 2、表 2的数据以及个体生长规律(李根前等,1999),可以将中国沙棘种群的生物量动态过程分为4个阶段:阶段Ⅰ:中国沙棘侵入裸地的最初7 a间,即中国沙棘灌草丛阶段,它依靠极强的根蘖能力和种子萌发使种群密度急剧增加,营养繁殖(地下部分)投资大。由于群落“过份拥挤”和分株之间的“根系连结”,“个体”通过迅速的树高生长来占领“制高点”以便“觅取”光资源,但径向生长量小。因此,这一阶段的生物量积累较少,估计值为4.914 4t·hm2,占上限的19.7%,平均生产量仅0.7021t·hm2a-1。阶段Ⅱ:中国沙棘无性系种群侵入立地的8~16 a期间(速增点出现在12 a),即幼林至成林阶段,在竞争中占有“空间优势”的“个体”,以迅速的径向生长扩大营养空间,使材积和生物量积累达到高峰。虽然种群密度因自疏而急剧下降,但“个体”生长的生物量积累大于密度下降引起的生物量减小。所以,该阶段的生产力最高,生物量估计值为15.171 2 t·hm2,占其上限的60.7%,平均生产量达1.685 7 t·hm2a-1a-1。阶段Ⅲ:在中国沙棘侵入立地16~20 a期间,“个体”开始衰老,生长量逐渐减小,生物量增长缓慢并趋于上限值。这一阶段的生物量积累估计值为3.628 1 t·hm2,占其上限的19.6%,平均生产量为0.907 0 t·hm2a-1。阶段Ⅳ:大约在中国沙棘种群侵入立地20 a以后,部分老龄林木枯死或倒伏,种群生物量开始下降。但在林窗中出现了幼苗,随着树冠的进一步疏开和幼苗生长,整个群落外貌呈幼树和中树与大树的异龄斑块镶嵌分布,种群生物量和密度又开始回升(图中样地C7)。
|
|

|
图 2 中国沙棘无性系种群生物量动态 Fig. 2 The biomass dynamics of clonal population of H.rhamnoides subsp. sinensis |
植物种群密度制约的核心是自然稀疏,即从群落郁闭开始,个体之间因竞争使种群个体数量不断减少从而保持群落的稳定性。植物种群的自然稀疏符合Yoda等提出的-3/2定律,即植物种群在自然稀疏过程中,其平均个体生物量与密度之间存在着幂函数关系,表达式为: W=C·D-a。式中,W为平均个体生物量; D为种群密度; C、a为模型系数。
由表 2可以看出,中国沙棘种群在由稳定到衰退的发展过程中,种群密度不断下降。因此,样地C3~C6的中国沙棘种群处于自然稀疏阶段,由于这一阶段每丛的蘖数变动在1.4~1.8之间,单轴型分株占60%以上,且合轴型分株主要出现在低龄级的分株中,因此,密度和生物量均以株或丛表示其近似值。据此回归得到中国沙棘种群的自然稀疏模型:W=2 272 292.973 D-1.559 8 (R=0.964 5, P<0.05)。式中,W为中国沙棘种群平均分株生物量(kg);D为密度(ind·hm2)。回归方程的幂指数非常接近-1.5,说明中国沙棘种群在自然稀疏过程中,种群密度和平均个体生物量之间的关系基本符合-3/2稀疏定律。这一结果说明,当把分株看成个体时,中国沙棘种群的密度调节符合稀疏定律。
-3/2稀疏定律说明,平均个体生物量每3个单位的变化对应2个单位的种群密度变化,即个体平均生物量的增长比由种群密度下降引起的种群生物量的下降要大。因此,中国沙棘种群生物量在自然稀疏过程中仍然保持着增长。
3 结论与建议中国沙棘种群密度表现出先增高后降低的趋势,并随着林窗的出现又开始回升,保证了中国沙棘种群在群落中的优势地位以及群落的稳定性。但是,与母株种群相比,子株种群的密度变化具有滞后效应,而且子株数量在种群中的比例越来越大,这样的稀疏速率差异使中国沙棘种群的基株多样性减小,即种群的组成由多数拥有较少分株的无性系向少数拥有较多分株的无性系转变。鉴于这种情况,适时适度地对中国沙棘种群进行人工干扰,如平茬、间伐等,可以提高群落的稳定性,有益于群落功能的持续发挥。同时,在林窗进行少量的种苗更新,对维持种群的基株多样性具有重要意义。
从种群增长到中衰阶段,中国沙棘种群生物量积累符合Logistic增长,速增期8~16a,速增点出现在12 a,此时已达数量成熟。此外,当把分株看成个体时,平均个体生物量和密度之间的关系符合-3/2幂法则。
董鸣. 1986. 缙云山马尾松种群数量动态初步研究. 植物生态学与地植物学学报, 10(4): 283-293. |
李根前, 黄宝龙, 唐德瑞, 等. 2001a. 毛乌素沙地中国沙棘无性系种群年龄结构动态与遗传后果. 应用生态学报, 12(3): 347-35O. |
李根前, 黄宝龙, 唐德瑞, 等. 2001b. 毛乌素沙地中国沙棘无性系生长调节. 应用生态学报, 12(5): 682-686. |
李根前, 黄宝龙, 唐德瑞, 等. 2001c. 毛乌素沙地中国沙棘无性系种群林缘扩散规律. 南京林业大学学报, 25(2): 9-13. |
李根前, 赵一庆, 唐德瑞, 等. 1999. 毛乌素沙地中国沙棘生长过程与水热条件的关系. 西南林学院学报, 14(1): 10-15. |
李秋元, 孟德顺. 1993. Logistic曲线的性质及其在植物生长分析中的应用. 西北林学院学报, 8(3): 81-86. |
刘庆, 钟章成. 1995. 无性系植物种群生态学研究进展及有关概念. 生态学杂志, 14(3): 40-45. |
刘庆, 钟章成. 1996. 斑苦竹无性系种群克隆生长格局动态的研究. 应用生态学报, 7(3): 240-244. |
朱志诚. 1992. 陕北黄土高原灌木林的类型及其动态特性. 陕西林业科技, (1): 36-42. |
Harper J L. 1981. Population biology of plants. New York: Academic Press, 637-643.
|
Hartnett D C, Bazzaz F A. 1985a. The genet and ramet population dynamics of Soliago canadensis in an abandoned field. Journal of Ecology, 73: 407-413. DOI:10.2307/2260483 |
Hartnett D C, Bazzaz F A. 1985b. The regulation of leaf, ramet and genet densities in experimental population of the rhizomatous perenial Solidago canadensis. Journal of Ecology, 73: 429-443. DOI:10.2307/2260485 |