 2004, Vol. 40
2004, Vol. 40文章信息
- 刘怀, 赵志模, 邓永学, 徐学勤, 李映平.
- Liu Huai, Zhao Zhimo, Deng Yongxue, Xu Xueqin, Li Yingping.
- 温度对竹盲走螨实验种群生长发育与繁殖的影响
- Temperature Effect on Development and Reproduction of Experimental Population of Typhlodrumus bambusae (Acari:Phytoseiidae)
- 林业科学, 2004, 40(1): 117-122.
- Scientia Silvae Sinicae, 2004, 40(1): 117-122.
-
文章历史
- 收稿日期:2002-03-26
-
作者相关文章


2. 四川省林业厅森防总站 成都 610061
2. Station of Forest Protection, Sichuan Forestry Bureau Chengdu 610081
毛竹(Phyllostachys heterocycla cv. pubescens)是我国南方最主要的经济树种,被林业部门定为退耕还林、保护环境、促进经济的主要森林树种(农业部,1998)。近年来,我国主要竹产区,如福建、浙江、四川等省的毛竹林出现大面积的竹叶黄化、落叶、甚至成片枯死的现象,各地区纷纷开展研究,从竹林老化、土壤衰退、大气污染、病虫发生等多个方面探讨枯死成灾的原因,最终认为是螨害,尤其是竹裂爪螨(Schizotetranychus bambusae)、食竹裂爪螨(S. celarius)等害螨类的危害是造成毛竹大面积枯死的主要因素。笔者于1998—2001在四川省国家级风景名胜区“蜀南竹海”开展了毛竹害螨发生与防治的试验研究,并在林间实际调查发现竹盲走螨(Typhlodromus bambusae)是毛竹林重要的捕食性天敌,对毛竹的一些害螨有较强的控制作用。但有关竹盲走螨生物学、生态学以及对害螨控制等方面研究的文献资料较少,国外一些学者对其生物学进行了初步研究(Mori et al., 1999;Yamamura et al., 1987;Saito, 1990)。在国内该螨为1996年报道的一个新纪录种(殷绥公等, 1996),对其研究仅见张艳璇等(2000)和Zhang Yanxuan等(1999)的报道。本试验在前人研究的基础上,以毛竹害螨优势种竹裂爪螨为食料,系统探讨温度对竹盲走螨生长、发育、繁殖的影响,以期为利用该螨进行毛竹害螨的生物防治奠定基础。
1 材料与方法 1.1 供试虫源在重庆北碚区缙云山毛竹林采集竹盲走螨,在室温条件下(温度25℃±0.5℃,RH70%~80%, L:D=12:12)用竹裂爪螨各螨态作为食料饲养若干代后作为供试螨源。竹裂爪螨的饲养采用植物叶片作为支持面的饲养方法(吴伟南等,1997),在室内常温下进行群集饲养,定期采集无病虫为害且颜色较绿的毛竹叶片作为饲料。
1.2 试验处理试验共设置16、20、24、28、32及36℃ 6种温度,以LRH-250-G型光照培养箱(广东医疗器械厂)控制温度,温差变幅均为±0.5℃,RH维持在70%~80%,每天给予12 h光照,采用植物叶片作为支持面的饲养方法进行饲养(吴伟南等,1997)。在直径为15 cm的培养皿内铺吸满水的塑料泡沫,其上铺一层吸水滤纸,取长13 cm左右,宽1 cm左右的毛竹叶片平铺于滤纸上,叶背朝上,并且用脱脂棉条压住叶片的叶尖、叶柄以及围住叶片四周, 然后移入竹裂爪螨雌成螨若干,让其自然产卵2~5 d,然后移去部分雌成螨,保留3~5头,使叶片上包含有竹裂爪螨卵、幼螨、若螨以及成螨等各螨态,以此作为饲养竹盲走螨的饲养皿。试验时,从群集饲养的竹盲走螨中,挑选若干头雌成螨于饲养皿的竹叶上,让其产卵,并每隔8 h检查1次,将8 h内所产卵视为同一时刻的初产卵,并将卵挑出,移入新的饲养皿叶片上,每叶片只挑入1粒卵,移至某一设置温度组内。进入实验时每温度组内的卵量为40~50粒。从移入卵时起,每隔8 h观察记录各处理的竹盲走螨的虫态变化、存活和死亡数、产卵量以及成螨寿命等。并根据各不同温度和螨态的情况,于每次检查时补若干竹裂爪螨的幼螨、若螨,保证每叶片上的食物包含有竹裂爪螨各螨态(主要以卵、幼螨和若螨为主)30~40头(粒),并根据不同温度每隔3~5 d更换1次含有竹裂爪螨新鲜竹叶。观察记录直到成螨死亡为止。
1.3 数据处理试验所得数据在计算机上用“SPSS”软件进行统计分析、模型拟合。方差分析(F测验)项目包括温度对竹盲走螨成虫寿命、产卵期、产卵量及存活率的影响,采用Duncan新复极差法进行多重比较。
2 结果与分析 2.1 不同温度条件下竹盲走螨各螨态发育历期竹盲走螨个体发育经过卵、幼螨、前若螨、后若螨和成螨5个阶段,在蜕皮前无明显的静息期。雄螨比雌螨发育稍快,先发育成熟的雄螨常滞留于雌螨左右,待雌成螨成熟后与之交配,交配型为“钝绥螨-盲走螨类型”(吴伟南等,1997)。不同温度下竹盲走螨的发育历期见表 1。
|
|
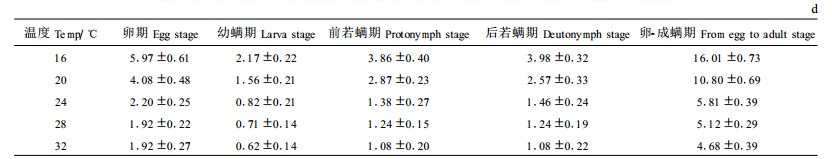
表 1说明,在16、20、24、28、32℃条件下,竹盲走螨卵至成螨历期分别为16.01、10.80、5.81、5.12和4.68 d。试验结果还表明,在36℃恒温条件下,竹盲走螨不能完成世代发育。在16℃~32℃范围内,卵期、幼若期以及世代历期都随着温度的升高而缩短,其中卵历期最长(5.97~1.92 d)、幼螨历期最短(2.17~0.62 d)。
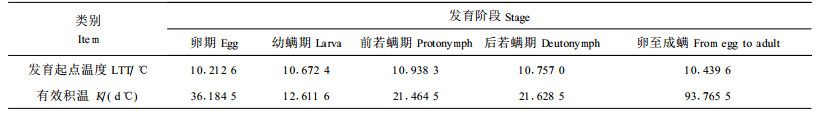
发育起点温度和有效积温是昆虫种群的基本生物学参数之一。本研究采用直接最优法计算各螨态的发育起点温度及有效积温(李典漠等1986),结果见表 2。从该表可以看出,竹盲走螨各螨态的发育起点温度约在10~11℃之间,其中卵至成螨的发育起点温度和有效积温分别为10.44℃和93.76d℃。
|
|
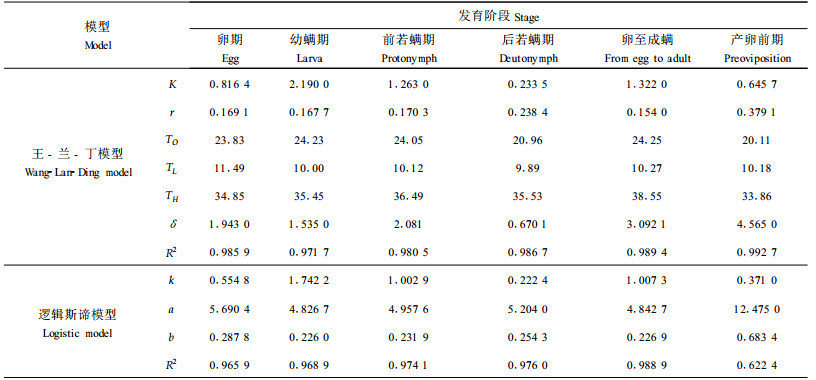
在昆虫(螨类)种群数量动态模拟中,一个很重要的模型是昆虫种群在各种温度条件下的发育率模型。根据表 1,将发育历期转换成发育速率(V)后,用Logistic模型(公式1)及王-兰-丁模型(公式2)拟合竹盲走螨卵、幼螨、前若螨、后若螨、卵至成螨以及产卵前期等各发育阶段的发育速率(V)与温度(T)之间的关系,求解出的模型参数见表 3,同时列出了各模型拟合效果的方差分析结果(用R2表示)。两种模型的公式分别为:
|
|
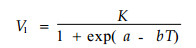
|
(1) |

|
(2) |
式(1)中,K:发育速率V的上限;T为温度;a、b为常数。式(2)中,K为潜在的饱和发育速率,等于最适发育速率V(TO)的二倍;r是发育速率随温度变化的指数增长率;TL、TH为最低和最高监界温度;TO为最适发育温度;δ为边界层宽度,其相对大小反映生物对极端温度的忍耐程度。
由表 3可见,竹盲走螨卵、幼螨、前若螨、后若螨以及卵至成螨的发育速率与温度关系的Logistic模型和王-兰-丁模型拟合效果基本一致;但王-兰-丁模型能更好地反映出竹盲走螨在高温下发育受到抑制的现象,其检验模型拟合的相关指数(R2)均大于Logistic模型的相关指数。并由王-兰-丁模型推知,竹盲走螨卵至成螨(未成熟阶段)的发育最低、最高临界温度分别为9.89℃与35.53℃,最适发育温度为20.96℃(表 3),这与本试验在36℃恒温条件下竹盲走螨不能完成世代发育的结果一致;各发育阶段的发育最低临界温度与有效积温法则计算出的发育起点温度也基本一致。
2.3 温度对竹盲走螨存活的影响实验表明,竹盲走螨各发育阶段的存活率与温度有着密切的关系。从表 4可以看出,在实验温度范围内(16~32℃),竹盲走螨各发育阶段的存活率均较高。温度对竹盲走螨幼螨的存活率几乎没有影响,其存活率均大于94%,20℃时最高,达100.00%,其原因可能是幼螨期发育历期短,对食物的要求较低,或者根本就不取食,可直接脱皮变成前若螨。从温度对竹盲走螨各发育阶段存活率的影响的还可以看出,卵期和前若螨期的对温度的变化表现得较为敏感。卵的孵化率在20℃和28℃条件下最高,均为97.50%,32℃条件下为85.00%;前若螨期存活率20℃时最高,为97.44%,16℃时较低,为82.86%。16、20、24、28、32℃各温度条件下卵至成螨的存活率分别为70.00%、92.50%、87.50%、85.00%和67.50%。
|
|

不同温度条件下竹盲走螨雌成螨的产卵前期、产卵期、产卵后期、寿命以及平均产卵量和每雌每日平均产卵量见表 5。
|
|
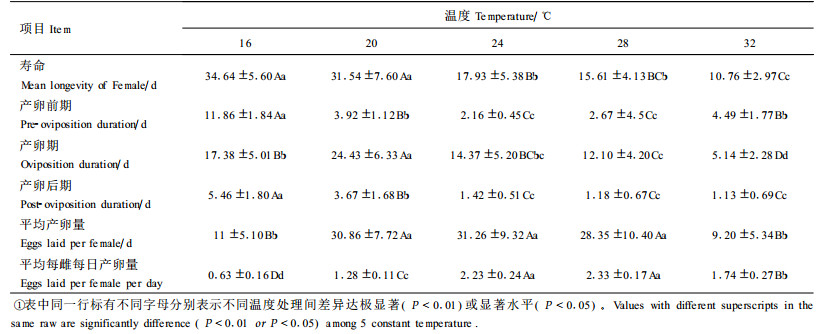
从表 5的结果可以看出,在70%~80%的相对湿度条件下,竹盲走螨雌成螨寿命在16℃条件下最长,平均为34.64 d,随着温度的升高,寿命逐渐缩短,32℃条件下最短,仅为10.76 d。在16℃低温条件下成螨的产卵前期和产卵后期较长,分别为11.86 d和5.46 d,是该温度下雌成螨寿命延长的主要原因。且适宜的温度范围内,产卵期和产卵前期随温度的升高而逐渐缩短,而超过某一温度后,其发育历期反而延长;在16~32℃范围内产卵后期随温度升高而缩短。
竹盲走螨后若螨蜕皮成为成螨后即可进行交配。在试验温度范围内,经过2.16~11.86 d后雌成螨开始产卵,由表 5可以看出,竹盲走螨每雌总产卵量在20℃与24℃ 2种温度条件下较高,分别为30.86粒和31.26粒,其次28℃时为28.35粒,但3种温度下产卵量的差异不显著。32℃条件下每雌平均总产卵量最低,仅为9.2粒。每雌每日平均产卵量表现为28℃最高,为2.33粒,而20℃与16℃分别为1.28和0.63粒,32℃下反而较高,为1.74粒,但由于32℃条件下产卵期最短,仅为5.14 d,是导致每雌总产卵量最低的直接原因。温度对竹盲走螨每雌平均总产卵量和每雌每日平均产卵量影响的变化趋势都呈开口向下的抛物线形式(如图 1、2)。因此,可用二次抛物线方程配合二者与温度(T)的关系,其方程为:平均产卵量(Y):Y=-171.694 2+17.273 2T-0.036 31T2(R=0.964 8* F(2.4)=36.379 6 P=0.026);平均每雌每日产卵量(Y):Y=-8.401 3+0.793 3T-0.014 8T2(R=0.970 6* F(2.4)=20.135 6 P=0.047)。

|
图 1 竹盲走螨每雌平均产卵量与温度的关系 Fig. 1 The relationship between eggs laid per female and temperature |

|
图 2 竹盲走螨每雌每日平均产卵量与温度的关系 Fig. 2 The relationgship between eggs laid per female per day and temperature |
由于不同恒温条件下的产卵期长短差异较大,平均每雌每日产卵量不能准确反映出温度变化对繁殖力所产生的影响。为了在同一水平上比较其繁殖力与温度的关系,现将不同温度下的产卵期划分成10个年龄级,具体划分情况如下,16℃条件下每3 d作为1个等级(产卵期最长为32 d),20℃每4 d作为1个等级(产卵期最长为41 d),24℃每3 d作为1个等级(产卵期最长为33 d),28℃每2 d作为1个等级(产卵期最长为19 d),32℃每1 d作为一个等级(产卵期最长为12 d),用每一等级内平均每雌日产卵量(Y)作为指标来衡量其繁殖力与温度的关系见图 3,其中:Y=N/(n·d)。式中N为每一等级内的总产卵量,n为雌螨数,d为等级间隔天数。

|
图 3 不同温度条件下竹盲走螨繁殖力与相对年龄等级的关系 Fig. 3 The relationship between fecundity and relative age level |
由图 3可以看出,当温度较低时(16℃),日产卵量少,生殖高峰不明显。20℃~28℃时,产卵由少到多,在第2年龄等级就可形成生殖高峰,其中24℃、28℃条件下产卵高峰期持续时间长,到第5年龄等级日平均产卵量才逐渐下降。32℃条件下,产卵期短,生殖高峰明显后移。
3 讨论在供试的温度(16~36℃)范围内,36℃条件下,竹盲走螨种群不能够完成世代发育,主要是因为卵在36℃的恒温条件下不能孵化。本试验将24℃条件下产生的初孵幼螨移入36℃恒温下进行饲养,其结果表现为各发育阶段存活率低,发育历期延长,即使有少量个体能发育至成螨,但都不能正常繁殖产卵,这与植绥螨的一些种类对温度的要求基本一致,如吴伟南等(1997)研究发现当温度超过32℃时,智利小植绥螨生长发育将受到抑制。
试验研究表明在16~32℃之间,随着温度升高,竹盲走螨的卵、幼螨、若螨的发育历期都逐渐缩短,亦即发育速率逐渐加快,但产卵前期在32℃延长。Saito对竹盲走螨的生物学进行过初步研究,结果表明在(25±0.5)℃的恒温条件下,以食竹裂爪螨为食时,卵到成螨的发育历期雄螨为6.1 d,雌螨为7.3 d(Saito,1990);而张艳璇等研究表明,在22~30℃的室温(变温)条件下以南京裂爪螨为食时,卵期为1.7 d,幼螨期为1 d,前若螨为0.8 d,后若螨0.8 d;本研究以竹裂爪螨为食时,(24±0.5)℃的恒温条件下,卵到成螨的发育历期5.81 d(张艳璇等,2000),形成差异的原因可能是不同的地理种群以及不同食料营养差异所致。
昆虫(螨类)发育速率与温度的关系是昆虫或螨类生物学研究的主要内容,它对于害虫害螨预测预报和益虫的利用有着巨大的价值,历来受到人们的重视。本研究分别用生态学中的基本模型Logistic方程和王-兰-丁模型拟合了竹盲走螨生长发育速率与温度的关系,其结果表明王-兰-丁模型能更好地反映不同温度条件下竹盲走螨生长发育情况,根据该模型推导出的各发育阶段对温度临界值与最适值与实验观察结果基本一致。
种群的死亡率(或存活率)、繁殖力、平均寿命是影响种群数量动态的基本生物学特征。温度作为一个重要的生态因子,作用于种群系统,必然引起上述基本生物学特征发生变化(赵志模,1984)。本试验在16~36℃的恒温条件下研究了温度对竹盲走螨存活、繁殖力以及平均寿命的影响。结果表明,竹盲走螨卵至成螨的存活率以20℃最高,达92.50%。各虫态比较而言,卵和前若螨期存活率对温度的变化表现得较为敏感,高温不利于卵的孵化,32℃时仅为85%,而其它温度处理均在92%以上。雌成螨的平均寿命在16℃最长,为34.64 d,随温度的升高,其寿命缩短。方差分析表明,16℃与24℃条件下的雌成螨寿命(分别为34.64和31.54 d),差异不明显。平均每雌总产卵量在20℃与24℃较高,分别为30.86粒·雌-1与31.26粒·雌-1。本文利用年龄等级法,以每一等级内平均每雌每日产卵量作为指标反映了种群增长和繁殖力特征与温度的关系,结果表明温度较低时(本试验在16℃),虽然雌成螨寿命与产卵期较长,但日产卵量少,不能产生明显的生殖高峰,32℃的较高温条件下,虽日产量较高,但产卵期短,日生殖高峰明显后移,即低温和高温均表现出不利于种群的增长。
李典谟, 王莽莽. 1986. 快速估计发育起点及有效积温法的研究. 昆虫知识, 23(4): 184-187. |
农业部汇编.中国农业年鉴.北京: 中国农业出版社, 1998
|
吴伟南, 梁来荣, 蓝文明.中国经济昆虫志.第五十三册.蜱螨亚纲.植绥螨科.北京: 科学出版社, 1997: 1-200
|
殷绥公, 余华星, 石纪茂, 等. 1996. 浙江省毛竹植绥螨1新种及1新记录(蜱螨亚纲:植绥螨科). 动物分类学报, 21(1): 58-61. |
张艳璇, 林坚贞, 刘巧云, 等. 2000. 毛竹叶螨的重要天敌竹盲走螨生态学研究. 昆虫天敌, 22(2): 54-58. DOI:10.3969/j.issn.1674-0858.2000.02.002 |
赵志模, 周新远. 1984. 生态学引论. 重庆: 科学技术文献出版社重庆分社, 1-65.
|
Mori K, Saito Y, Sakagami T. 1999. Effects of the nest web and female attendance on survival of young in the subsocial spider mite Schizotetranychus longus (Acari:Tetranychidae). Experimental and Applied Acarology, 23(5): 411-418. DOI:10.1023/A:1006165606428 |
Saito Y. 1990. Life history and feeding habit of T. bambusae, a specific predator of S. celarius (Acari:phytoseiidae; Tetranychidae). Experimental and Applied Acarology, 10(1): 45-51. DOI:10.1007/BF01193972 |
Yamamura N, Saito Y. 1987. Biparental defence in a subsocial spider mite. Trends in Ecology and Evolution: 261-262. |
Zhang Y X, Zhang Z Q, Liu Q Y, et al. 1999. Biology of Typhlodromus bambusae (Acari:Phytoseiidae), a predator of Schizotetranychus nanjinqensis (Acari:Tetranychidae) injurious to bamboo in Fujian, china. Systematic and Applied Acarology, 4: 57-62. DOI:10.11158/saa.4.1.8 |