 2003, Vol. 39
2003, Vol. 39文章信息
- 叶功富, 张清海, 林益明, 吴锡麟.
- Ye Gongfu, Zhang Qinghai, Lin Yiming, Wu Xilin.
- 沿海防护林生态系统不同群落生物量和能量的研究
- STUDY ON BIOMASS AND ENERGY OF DIFFERENT COMMUNITIES ON THE COASTAL FOREST ECOSYSTEM
- 林业科学, 2003, 39(专刊1): 8-14.
- Scientia Silvae Sinicae, 2003, 39(专刊1): 8-14.
-
文章历史
- 收稿日期:2002-12-20
-
作者相关文章


2. 福建省农林大学林学院 南平 353001;
3. 厦门大学生命科学学院 厦门 361005
2. Forestry College of Fujian Agriculture and Forestry University Nanping 353001;
3. School of Life Sciences, Xiamen University Xiamen 361005
沿海防护林是海岸主要的生态屏障, 以木麻黄(Casuarina equisetifolia)为主的生态系统在防风护沙、改善沿海的生态环境和促进经济建设等方面有着难以替代的作用(叶功富等, 2000)。本文通过对基干林带不同群落现存量生物量、各组分的热值、能量现存量、能量分布和能量归还量的研究, 深入了解了沿海基干防护林带生态系统的结构与功能, 为沿海防护林的更新改造, 实现可持续经营提供理论依据。
1 试验地概况试验地设在福建省东山县国有防护林场, 位于福建省南部, 东经117°18′, 北纬23°40′。属亚热带海洋性季风气候, 年平均气温20.8 ℃, 极端最高气温36.6 ℃, 极端最低气温3.8 ℃, 全年无积雪, 无霜冻, 年均降水1 164 mm, 年均蒸发2 028 mm, 全年干湿季节明显, 每年的11月至翌年的2月为旱季, 大部分的降水集中于台风多发的5 —9月, 年均台风5.1次。土壤以沙土为主, 整个生态系统以木麻黄为主, 自从1992年开始对海岸防护林进行多树种的引种。本次研究的样地选择1992年造林的木麻黄-厚荚相思(Acasia crassicarpa)群落, 由于厚荚相思后期生长较快, 林分郁闭后, 木麻黄受到了厚荚相思排挤成为下木; 木麻黄-湿地松(Pinus elliottii)群落, 群落中两者均为阳性树种, 彼此之间的生长势较为相似, 两者的生长情况接近; 木麻黄群落样地内无灌木, 草本稀少。
2 研究方法 2.1 生物量测定在木麻黄-厚荚相思、木麻黄-湿地松和单一木麻黄群落中各设立20 m×20m样地3块, 对样地内的林木进行每木检尺和树高测定, 根据每木检尺的胸径和树高值, 选取平均木, 按2 m区分段法进行树干解析, 枝叶部分采用分层收获法, 地下部分采用壕沟法。称取各组分的鲜重, 并取样带回实验室, 在80 ℃下烘干待测, 另取小样在105 ℃下烘干, 求含水率。根据各组分的干湿比, 求各组分的生物量。林下无灌木, 草本稀少, 故本研究中不予考虑。
2.2 凋落物收集2002年在木麻黄-厚荚相思、木麻黄-湿地松和木麻黄群落样地中各设立5个1 m ×1 m收集筐, 网孔径为0.5 mm。每月收集1次, 样品带回实验室, 在80 ℃下烘干, 称重, 各月累加得到凋落物的量。
2.3 热值测定样品烘干后, 磨粉处理过6号筛贮存备用。用热量计法测定热值, 仪器采用长沙仪器厂生产的GR -3500型微电脑氧弹式热量计。样品热值以干重热值(每克干物质在完全燃烧条件下所释放的总热量)和去灰分热值表示。测定环境是在空调控温20 ℃左右, 每样品重复2 ~ 3次, 误差控制在±0.20 kJ, 每次试验对仪器用苯甲酸标定。
灰分测量采用干灰化法, 样品在马福炉550 ℃下灰化5 h后测定灰分含量。去灰分热值=干重热值(1 -灰分含量)。去灰分热值除去灰分含量不同的干扰, 更能够反映植物体各组分热值情况。
3 结果与分析 3.1 群落生物量及凋落物归还量 3.1.1 群落现存生物量和分布生物量是指在群落中一定时间内单位面积上的植物量, 是生态系统的结构指标之一, 是生态系统功能的物质基础和食物链的能流基础(彭少麟等, 1998)。2002年11月, 对木麻黄-厚荚相思、木麻黄-湿地松和木麻黄群落的生物量进行了测定, 群落生物量和各组分生物量见表 1。
|
|
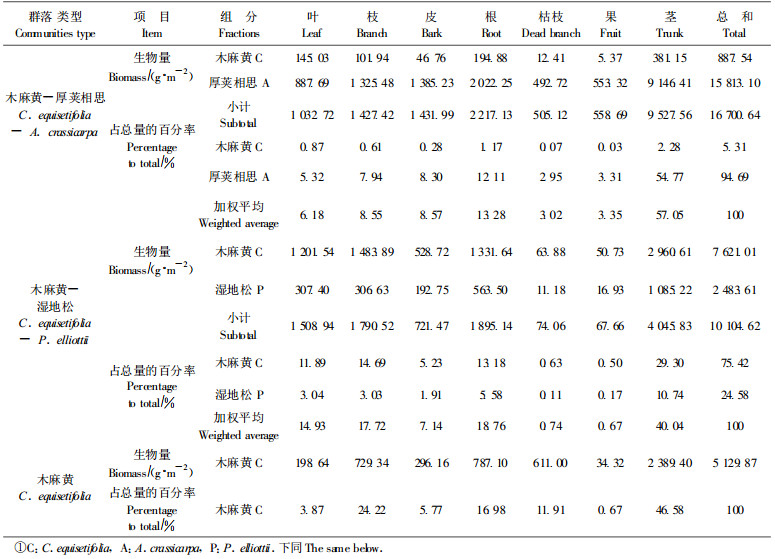
从表 1可见, 沿海防护林生态系统中不同的群落结构在生物量上有着较大的差异, 木麻黄-厚荚相思群落的生物量最大为16 700.64 g·m-2, 高于南亚热带鹤山豆科混交林11 080 g·m-2、马占相思(Acacia mangium)林12 423 g·m-2和针叶混交林7 430 g·m-2(彭少麟等, 1998), 也高于湖南会同县杉木(Cunninghamia lanceolata)林15 631 g·m-2(冯宗炜等, 1982)和木麻黄林7 965 g·m-2(叶功富等, 1996), 低于南亚热带黑石顶天然林35 798.00 g·m-2(彭少麟等, 1998), 中亚热带武夷山甜槠(Casfanopsis eyrei)林40 730 g· m-2(林益明等, 2001)和武夷山黄山松(Pinus taiwanensis)群落19 128.0 g·m-2(林鹏等, 1999), 群落中木麻黄887.54 g·m-2, 占群落总生物量的5.31%, 厚荚相思15 813.10 g·m-2, 占群落总生物量的94.69%, 在群落中各组分的生物量以茎最大, 9 527.56 g·m-2, 占总量的57.05%, 枯枝最小, 505.12 g·m-2, 占总量的3.02%, 群落中各组分含量的顺序为:茎>根>皮>枝>叶>果>枯枝; 木麻黄-湿地松群落其次, 总生物量为10 104.62 g·m-2, 略低于南亚热带鹤山湿地松林10 226 g·m-2和马尾松(Pinus massoniana)林10 877 g·m-2(彭少麟, 1998), 高于南亚热带鹤山针叶混交林7 430 g·m-2(彭少麟等, 1998), 低于中亚热带武夷山黄山松林19 128.0 g·m-2 (林鹏等, 1999), 高于湖南会同马尾松林10 000 g·m-2 (冯宗炜等, 1982), 群落中木麻黄为7 621.01 g·m-2, 占总量的75.42%, 湿地松为2 483.61 g·m-2, 占总量的24.58%, 群落内各组分中, 茎生物量最大, 4 045.83 g·m-2, 占群落总生物量的40.04%, 果最小, 67.66 g·m-2, 占群落总生物量的0.67%。群落中各组分顺序为:茎>根>枝>叶>皮>枯枝>果; 单一木麻黄群落最小, 总生物量为5 129.87 g·m-2, 高于南亚热带小良桉树(Eucalyptus)林4 200 g·m-2(彭少麟等, 1998), 低于同龄木麻黄林7 965 g·m-2(叶功富等, 1996), 其中茎的生物量最大2 389.40 g·m-2, 占总生物量的46.58%, 果最小34.32 g·m-2, 占总生物量的0.67%, 群落中个组分顺序为:茎>枝>根>枯枝>皮>叶>果。
从表 1可知, 木麻黄-厚荚相思群落生物量16 700.64 g·m-2, 是单一木麻黄群落5 129.87 g·m-2的3.26倍, 是木麻黄-湿地松群落1 010.62 g·m-2的1.65倍, 木麻黄-湿地松群落是单一木麻黄群落的1.97倍。生物量是森林生态系统发挥功能的物质基础, 混交防护林群落有更高的生物量, 即混交群落对环境资源有更高的利用率、更高的生产力和更好的生态功能, 起着更高的防护作用。
3.1.2 凋落物归还量森林群落中以凋落物的形式向土壤归还物质和能量, 为土壤中的生命活动提供能源和维持生态系统功能正常运行起着重要的作用。通过对木麻黄-厚荚相思、木麻黄-湿地松和木麻黄群落凋落物的收集、调查可知, 在不同的群落中, 凋落物的归还量存在着一定的差异, 木麻黄-厚荚相思群落的最大为1 497 g·m-2, 其次木麻黄-湿地松群落为1 417.77 g·m-2, 单一木麻黄群落最小为95.10 g·m-2, 从表 1可知3个群落的生物量, 凋落物与现存生物量成正相关。
3.2 群落能量现存量 3.2.1 群落中各组分热值群落中各组分热值是计算群落能量现存量的基础, 根据群落中各组分的生物量, 可推算群落能量的现存量。从表 2可见:木麻黄-厚荚相思群落中:木麻黄各组分干重热值为19.83 ~ 21.52 kJ·g-1, 各组分的顺序是:叶>树皮>根>枯枝>果>茎>枝; 灰分含量0.51%~ 3.53%, 以果为最大, 枯枝最小; 去灰分热值20.02 ~ 22.31 kJ·g-1, 其各组分顺序是:叶>皮>根>果>枯枝>枝>茎。厚荚相思各组分干重热值为20.00 ~ 23.77 kJ·g-1, 其各组分顺序是:叶>茎=根>枯枝>枝>果>皮; 灰分含量0.68%~ 3.57%, 其中叶最大, 枯枝最小; 去灰分热值20.23 ~ 24.65 kJ·g-1, 各组分顺序是:叶>根>枝>茎>枯枝>果>皮。
|
|

在木麻黄-湿地松群落中, 木麻黄各组分干重热值19.47 ~ 20.83 kJ·g-1, 各组分顺序是:叶>根>果>枝>皮=茎>枯枝; 灰分含量0.68%~ 5.58%, 叶最大5.58%, 茎最小0.68%;去灰分热值20.00 ~ 22.06 kJ·g-1, 各组分顺序是:叶>果>根>皮>枝>茎>枯枝。湿地松各组分干重热值20.54 ~ 21.54 kJ·g-1, 各组分顺序是:枯枝>叶>枝>茎>根>皮>果; 灰分含量0.85%~ 2.35%, 根含量最大2.35%, 茎最小0.85%;去灰分热值20.93 ~ 21.92 kJ·g-1, 叶>枯枝>枝>根>茎>皮>果。
单一木麻黄群落中, 各组分干重热值19.98 ~ 21.27 kJ·g-1, 各组分顺序是:枯枝>皮>叶>茎>果>枝>根。植物组分热值及其器官热值差异主要是受自身组成、结构和功能的影响, 此外还受光照强度、日照长短(林益明等, 2001)。群落中植物体平均热值为:灰分含量0.42%~ 5.51%, 果最大5.51%, 茎最小0.42%;去灰分热值20.28 ~ 21.44 kJ·g-1, 各组分的顺序是:果>枯枝>叶>皮>茎>根>枝。
从上述可知, 木麻黄各组分干重热值在19.47 ~ 21.52 kJ·g-1, 植物体平均热值为20.52 kJ·g-1, 高于武夷山甜槠林的17.06 ~ 20.28 kJ·g-1, 植物体平均热值19.17 kJ·g-1(林益明等, 2001), 也高于长白落叶松(Larix gmelinii)林的18.52 ~ 19.85 kJ·g-1, 植物体平均热值18.97 kJ·g-1(刘世荣等, 1992); 厚荚相思各组分干重热值20.00 ~ 23.77 kJ·g-1, 植物体平均热值为20.47 kJ·g-1, 同样高于武夷山甜槠林, 也高于武夷山黄山松的16.27 ~ 22.14 kJ·g-1, 植物体平均热值19.86 kJ·g-1(林鹏等, 1999)。湿地松各组分干重热值20.54 ~ 21.54 kJ·g-1, 植物体平均热值为21.30 kJ·g-1, 高于长白落叶松与武夷山黄山松的相应热值。不同组分中一般叶的热值较大, 茎、枝和根的热值较小。上述出现枯枝大于活枝, 这与Ovington(1961)研究欧洲赤松(Pinus sylvestris)林发现11 a生的欧洲赤松枯枝热值大于活枝相同。植物组分热值及其器官热值差异主要是受自身组成、结构和功能的影响, 此外还受光照强度、日照长短(林益明等, 2001)。木麻黄在不同群落中热值不同的主要是不同群落的光照强度不同。从上述可知, 沿海防护林中树种各组分热值较高, 各树种光能利用率较高的树种。
3.2.2 群落中能量现存量能量值的测定能够更好地评价物质在生态系统中, 各成分间转移过程中质和量的变化规律, 应用能量概念研究植物群落更能够反映出群落对自然资源的利用情况。根据群落中各组分的热值及其相应的生物量计算而得, 各组分热值见表 2, 各组分的生物量见表 1。从表 3可见:木麻黄-厚荚相思群落能量现存量为341 685.70 kJ·m-2, 高于南亚热带鹤山大叶相思(A. auriculaeformis)林的172 200 kJ·m-2和马占相思林的278 200 kJ·m-2, 低于南亚热带鼎湖山针阔混交林的739 960 kJ·g-1 (彭少麟等, 1998)。群落中木麻黄为18 033.30 kJ·m-2, 占总量的5.28%, 厚荚相思为323 652.39 kJ·m-2, 占总量的94.72%;茎最大, 占总量的56.61%, 枯枝最小, 占总量的2.93%, 各组分顺序为:茎>根>皮>枝>叶>果>枯枝。木麻黄-湿地松群落能量现存量210 128.30 kJ·m-2, 高于南亚热带鹤山针叶混交林的132 000 kJ·m-2(彭少麟等, 1998), 低于中亚热带武夷山黄山松林的379 832.3 kJ·m-2; 群落中木麻黄157 222.20 kJ·m-2, 占总量的74.82%, 湿地松54 906.10 kJ·m-2, 占总量的25.18%;各组分中茎最大, 占总量的39.24%, 果最小, 占总量的0.68%, 群落中各组分的顺序是:茎>根>枝>叶>皮>枯枝>果。单一木麻黄群落能量现存量105 751.40 kJ·m-2, 茎最大, 占总量的46.93%, 果最小, 占总量的0.66%, 各组分顺序是:茎>根>枝>枯枝>皮>叶>果。
|
|
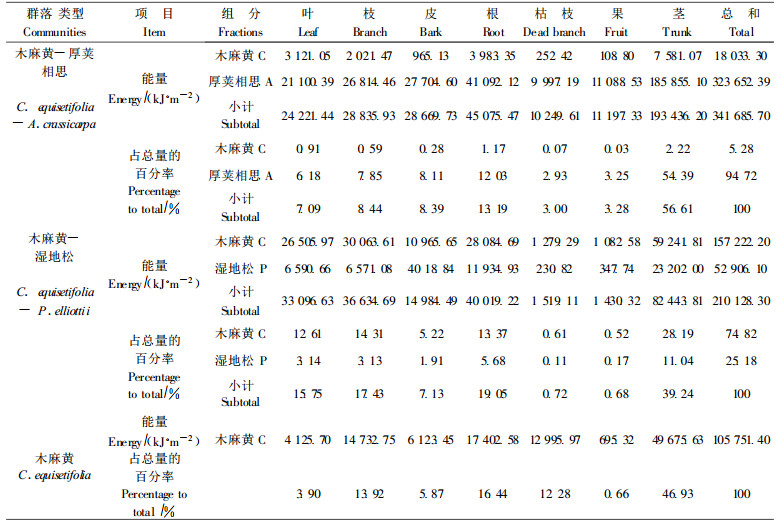
从上述可知, 木麻黄-厚荚相思群落的能量现存量最大, 是木麻黄-湿地松群落的1.63倍, 是单一木麻黄群落的3.23倍。木麻黄-湿地松群落能量现存量是单一木麻黄群落的1.99倍。可见复合群落的能量现存量要高于单一群落的能量现存量; 在复合群落中木麻黄-厚荚相思群落能量现存量最大。能量现存量的积累是森林生态系统生长、发育和演替的结果, 只有能量现存量达到一定水平, 形成较强的能量贮存和较强的能流, 才会形成较多的空间结构, 群落才具有更强的抗逆性。沿海防护林生态系统中营造复合生态系统有高的能量现存量和强的抗逆性, 能更好地发挥防护作用。
3.3 群落能量归还量生态系统中每年以枯枝落叶的形式向土壤归还养分和能源, 为林木的生长提供必要的养分, 同时也作为土壤腐生生命活动的能源, 以维持生态系统功能的正常运行(刘世荣等, 1992)。群落能量的归还量根据群落凋落物的归还量和凋落物干重热值计算而得。从表 4可见, 木麻黄-湿地松群落的能量归还量最大, 为30 595.48 kJ·m-2, 木麻黄-厚荚相思群落的能量归还量其次为28 728.20 kJ·m-2, 木麻黄最小为2 061.77 kJ·m-2。木麻黄-湿地松群落能量的归还量是木麻黄-厚荚相思群落的1.06倍, 是单一木麻黄群落的14.84倍, 木麻黄-厚荚相思群落是单一木麻黄群落的13.93倍, 可见复杂群落的凋落物能量归还量大, 有利于土壤生命活动, 改良土壤, 有利于群落中物种的生长, 使群落结构和功能更趋向于稳定。
|
|

在不同的群落结构中, 现存生物量上存在着一定的差异, 总体上复合群落的生物量要高于单一群落的生物量, 在复合群落中以木麻黄-厚荚相思群落的最大, 为16 700.64 g·m-2, 木麻黄-湿地松群落次之, 10 104.62 g·m-2, 单一木麻黄群落为5 129.87 g·m-2。复合群落有较高的生物量现存量。
在不同的群落中各组分干重热值情况是:在木麻黄与厚荚相思群落中, 木麻黄各组分干重热值19.83 ~ 21.52 kJ·g-1, 植物体平均热值为20.32 kJ·g-1, 厚荚相思各组分干重热值20.00 ~ 23.77 kJ·g-1, 植物体平均热值为20.47 kJ·g-1, 整个群落植物体的干重热值平均为20.46 kJ·g-1; 在木麻黄-湿地松群落中, 木麻黄各组分热值19.47 ~ 20.83 kJ·g-1, 植物体平均热值为20.63 kJ·g-1, 湿地松各组分热值20.54 ~ 21.54 kJ·g-1, 植物体平均热值为21.30 kJ·g-1, 整个群落植物体平均热值20.80 kJ·g-1; 单一木麻黄群落各组分热值19.98 ~ 21.27 kJ·g-1, 整个群落植物体平均热值20.61 kJ·g-1。总体上, 沿海防护林树种的植物体热值, 具有较高的光能利用率, 是生长较迅速的树种。
在不同群落能量存在着一定的差异, 木麻黄-厚荚相思群落中能量现存量为341 685.70 kJ·m-2, 木麻黄-湿地松群落中能量现存量为210 128.30 kJ·m-2, 单一木麻黄群落能量现存量为105 751.40 kJ·m-2。总体上复合群落的生物量和能量均要高于单一树种的群落, 木麻黄-厚荚相思群落中的生物量和能量分别是木麻黄-湿地松群落的1.65倍和1.63倍, 是单一木麻黄群落的3.26倍和3.23倍, 木麻黄-湿地松群落的现存生物量和能量分别是单一木麻黄群落的1.97倍和1.99倍。可见在沿海防护林生态系统建设中应以混交林为主, 使沿海防护林体系有更稳定的结构和更高的防护效果。
冯宗炜, 陈楚莹, 张家武, 等. 1982. 湖南会同县两个森林群落生产力. 植物生态学与地植物学学报, 6(4): 257-266. |
林鹏, 林益明, 李振基, 等. 1999. 武夷山黄山松群落能量的研究. 生态学报, 19(4): 504-507. DOI:10.3321/j.issn:1000-0933.1999.04.013 |
林益明, 杨志伟, 李振基. 2001. 武夷山常绿林研究. 厦门: 厦门大学出版社, 42-48;58-63;124-127.
|
刘世荣, 王文章. 1992. 落叶松人工林生态系统净初级生产力形成过程中的能量特征. 植物生态学与地植物学学报, 16(3): 209-219. |
彭少麟, 任海. 1998. 南亚热带森林生态系统的能量生态研究. 北京: 气象出版社, 45-60;74-79.
|
叶功富, 林银森, 吴寿德, 等. 1996. 木麻黄林生产力动态变化的研究. 防护林科技, (专): 17-20. |
叶功富, 谭芳林, 徐俊森, 等. 2000. 木麻黄基干林带防风效应及其与林带结构关系的研究. 防护林科技, (专): 103-107. |
Ovington J D. 1961. Some aspects of energy flow in plantations of Pinus sylvestris. Annuals of Botany, 15(27): 12-20. |