 2003, Vol. 39
2003, Vol. 39文章信息
- 刘金福, 于玲, 洪伟, 李俊清, 许忠实.
- Liu Jinfu, Yu Ling, Hong Wei, Li Junqing, Xu Zhongshi.
- 格氏栲林林窗物种多样性动态规律的研究
- STUDY ON DYNAMIC PATTERN OF SPECIES DIVERSITY IN GAPS OF CASTANOPSIS KAWAKAMII FOREST
- 林业科学, 2003, 39(6): 159-164.
- Scientia Silvae Sinicae, 2003, 39(6): 159-164.
-
文章历史
- 收稿日期:2002-07-22
-
作者相关文章



2. 国家林业局科技发展中心 北京 100714;
3. 北京林业大学资源与环境学院 北京 100083
2. Technology Development Center of National Forestry Bureau Beijing 100714;
3. College of Resources and Environment, Beijing Forestry University Beijing 100083
格氏栲(Castanopsis kawakamii),也称青钩栲,属国家二级保护植物,是壳斗科常绿阔叶大乔木,为亚热带珍贵稀有树种之一。自然分布仅限福建、江西、广东和广西等地,多零星生长在海拔200~1 000 m丘陵地带的常绿阔叶林中。在福建三明小湖地区有近700 hm2的林分,多由百年以上的大树组成,是中国较为罕见的天然群落,有其独特的自然景观(林鹏等,1986;樊后保,2000)。有关学者对格氏栲林开展了一些研究(刘金福等,1998;1999a;1999b;2001b;2001a;2001c;2002a;2002b;2002c),但有关格氏栲林多样性动态研究并不多见。林窗的大小与发育阶段是林窗干扰状况的两个重要参数,通过对物种多样性与林窗大小发育阶段的研究,结合植物群落的群落生态学特征进行系统分析,有利于了解格氏栲树冠干扰与格氏栲林生物多样性维持的机理,为珍稀格氏栲林的合理保护与可持续经营提供科学依据。
1 研究区自然概况格氏栲林位于福建省三明市西南方,面积近700 hm2,地处于26°7′~26°10′N、117°24′~117°27′E,属福建武夷山东伸支脉地带,东南方为戴云山脉,海拔200~500 m,多地形雨,年均温度为19.4℃,极端最低温为-5.5℃,最高温为40℃,大于10℃年积温为6 215℃;平均降雨量1 500 mm,3—8月的降雨量为全年的75%,大于0.1 mm的年降雨日数163.3 d;平均相对湿度79%;年平均风速1.6 m·s-1;四季分明,春夏季温热多雨,秋冬稍干旱,表现出温热湿润亚热带气候特色,有利于植物生长、岩石风化、土壤淋溶作用和富铝化作用。格氏栲林下土壤类型主要为暗红壤,其次为紫色土,土层比较厚,多数达1.0~1.5 m,土层腐殖丰富,水肥条件较好,植物种类丰富,郁闭度多达0.8左右,林木树干通直,但分枝较多,树皮有一定的剥裂,林相整齐,整个群落浓郁苍翠,形成中亚热带常绿阔叶树所特有的外貌特征,由于保护区海拔低,无明显垂直分布带,格氏栲多数分布海拔450 m以下(刘金福等,1998;1999a)。由于调查区位于格氏栲自然保护区管理站东面,属保护区范围内,很少受人为破坏,基本上保持天然次生林特征,其调查区海拔高度为200~405 m,植被是中亚热带低山常绿阔叶林,以格氏栲为群落的优势种。
2 研究方法 2.1 调查方法根据对格氏栲天然林踏查基础上,采用样线调查法(Runkle,1982;1985;臧润国等,1998),从格氏栲自然保护区管理站左侧林缘向森林走约500 m,以某一随机点开始调查,从起点沿罗盘仪所指的方向,由西向东方穿行,遇林窗则停下,共调查40个林窗样地,做下述记载:(1)测定扩展林窗内的长轴和与之垂直的短轴,考虑所调查的林窗皆接近于椭圆形,林窗面积一般按椭圆来计算(Runkle,1982);(2)测定形成年龄,先进行对林窗内形成木进行每木检尺,采用解析木资料或文献(刘金福等,1999b)等手段测定立枯或倒木的树龄,运用追溯法推测出林窗的形成年龄(洪伟等,2000);(3)所确定的每个林窗,记载其环境特征和周围群落性质,对构成林窗边界的各大乔木进行每木调查,采用样方法取样,在每个林窗内中心区同样位置设立2个5 m×5 m小样方(考虑有小于80 m2的林窗),并在离每个林窗边缘10 m处随机设置1个5 m×5 m的非林窗对照区样方,分别记载林窗内及非林窗林分的的乔木、灌木的种类、个体数、高度、冠幅,及各样方中草本、乔灌幼苗的种类、个体数和盖度等内容。
2.2 数据处理以研究区域40个林窗和40个非林窗对照样方中的物种个体数为基础,根据调查的林窗面积大小和年龄分布状况,将林窗按面积大小和年龄分别分成为6个等级,进行归类统计,各等级标志值及其林窗个数分布情况见表 1、2,应用下列多样性测定指标对不同级内的每个林窗各层次的物种多样性进行测度,然后将不同等级内的所有林窗不同层次的多样性指数分别进行加权平均,即得到各级内平均每个林窗的各层次多样性指数值,以此为基础进行各级林窗的物种多样性对照与分析。
|
|
|
|
采用5个目前应用较为普遍的计算公式,测度格氏栲林林窗物种多样性指标(吴承祯等,1996;臧润国等,1999;洪伟等,1999;刘金福等,2002a)。
丰富度指数:Margalef指数R=(S-1)/ln N,其中:S为物种总数即丰富度(下同);N为观察到的个体总数(随样本规模的增大而增大)。
物种多样性:多样性指数是物种丰富度与相对多度综合而成。Shannon-Wiener指数:H= -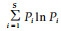
群落均匀度:指群落中物种的多度分布的均匀程度。Pielou指数Js=H/lnS,式中:S为物种总数;H为Shannon-Wiener指数。
生态优势度:反映群落逐种群优势状况的指标。Simpson指数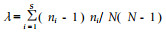
均优多指数:指综合物种丰富度、均匀度和生态优势度来表示群落多样性指数的大小:Z=(Js-λ)×S,式中:Js、λ、S意义同上。
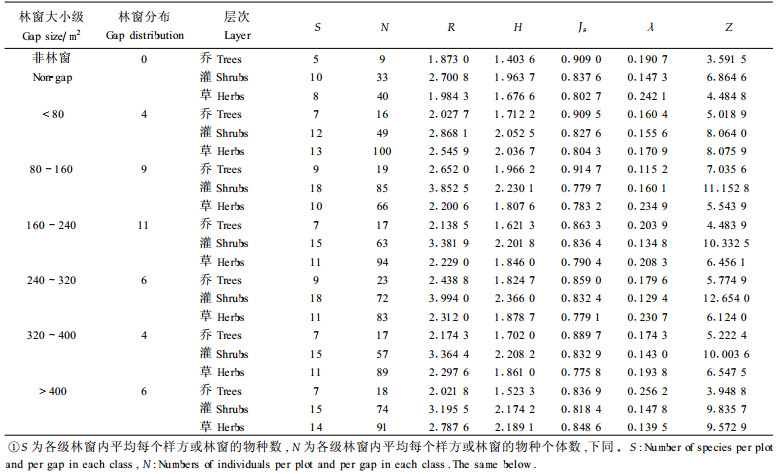
3 结果分析 3.1 不同大小林窗内的物种多样性变化采用上述5个公式计算不同大小级林窗中各层次的多样性指标(见表 1)。经方差分析检验,6种大小林窗、非林窗林分的多样性测度存在明显差异(p < 0.05)。可见,不同大小林窗的生态微环境不同,其中更新植物的种类和数量也发生相应的变化,表现出物种多样性特征明显产生差异。同样大小林窗内,由于不同种类对环境的适应要求不一样,不同层次的多样性也表现差异。就林窗内乔木层而言,物种丰富度指数R、物种多样性指数H、均匀度指数Js的变化趋势与均优多指数Z相一致,均随着林窗面积增大的变化曲线呈现双峰型,在80~160 m2的林窗内达到最大,即该大小林窗的物种多样性最大,随着林窗面积进一步扩大,R、H、Z均在240~320 m2的林窗中又出现一个峰值,Js在320~400 m的林窗中出现另一个峰值,但R、H、Js、Z等指数均明显表现出略有下降趋势,均在大于400 m2的林窗中为较小,而R、H、Z在非林窗林分中为最小,Js在小于160 m2的林窗内比非林窗林分大,主要在于格氏栲林上层以格氏栲等优势种所构成的,非林窗林分个体数量分化程度略有强烈。个体生态优势度λ变化趋势几乎与均匀度相反,在80~160 m2的林窗内为最小,表明80~160 m2的林窗的乔木层的植物由多物种组成,优势种并不突出,随着林窗形成面积扩大,林窗内个体数量分化程度逐步由弱逐步变强,优势种越来越来明显,大于400 m2的林窗的优势种表现最为明显,与面积较大的林窗给喜光物种优势种格氏栲、木荷(Schima superba)创造良好环境有关。
对林窗灌木层而言,其物种丰富度指数R、物种多样性指数H和均优多指数Z变化趋势基本与物种数S相似,均呈双峰型,且变化趋势大致相似,尤其H与Z变化趋势基本一致。表现为R、H、Z的最大峰值均在240~320 m2的林窗中,均在形成面积不大的林窗(小于80 m2)中、非林窗林分中为最小,在80~160 m2的林窗中出现另一个峰值。均匀度指数Js的变化曲线则与R、H、Z有所差异,在80~160 m2的林窗中为最小,在非林窗林分中、160~240 m2的林窗中达到较大,160~400 m2的林窗中均匀度变化不大。生态优势度λ几乎与均匀度指数相反,在240~320 m2的林窗内为最小,在80~160 m2林窗内物种优势特别明显,表明林窗灌木层在160~400 m2林窗内物种组成最为复杂且各种类株数分布比较均匀,优势种并不十分突出,继续扩大林窗面积,呈现出物种多样性有所下降、优势种明显突出的趋势。整体上看,林窗内灌木层的物种数多、丰富程度高,但各级林窗内灌木层个体数在不同物种之间的均匀分布低于非林窗林分,且大于160 m2的林窗内的个体生态优势度均小于非林窗林分,主要与灌木层处于更新层处,该层物种利用林窗的能力差异较大以及物种大多在林窗形成后生长和发育的有关。
对林窗草本层而言,物种丰富度指数R、物种多样性指数H、均匀度Js、均优多指数Z的变化曲线与物种数S一样均呈凹形,即中间低,两端高。这些指数均在小于80 m2的林窗内为较大,在大于400 m2的林窗内达到最大,该大小林窗草本层有较高的物种多样性,株数分布较为均匀,而R、H、Z在80~160m2林窗内、非林窗林分中为最小,Js则在320~400 m2的林窗内为最小;生态优势度值λ与多样性指数相反,各级林窗个体优势度均小于非林窗林分,在大于400 m2林窗中为最小,可见林窗内草本层个体数量分化强烈程度也低于非林窗林分。草本层的表现形式可能与下列原因有关:林窗不大时,环境变化对草本层物种多样性、均匀度影响并不大,保留林冠下原有物种,且增加一些喜光物种,增大物种多样性;随着林窗面积增大,受到光反射等因子影响越来越大,耐荫性物种可能会逐渐消失,导致多样性减小;林窗面积进一步扩大,可能侵入较多物种而增加其丰富度,因而出现较高的物种多样性。
总体上看,林窗内乔、灌木层的物种丰富度指数R、物种多样性指数H和均优多指数Z变化趋势随着林窗大小呈波浪状起伏,略有下降趋势,表明乔、灌木层物种多样性并不随着林窗面积增大而增多,而且面积过大或过小的林窗内的物种均不可能拥有最大的多样性,可能与林窗内的生态因子的变化有关(安树青等,1997),太小的林窗内光照较弱,不利于耐荫性较小树种的生存和发展;太大的林窗内光照较强,不利于耐荫性较大树种的生存和发展,可见,格氏栲林林窗达到一定大小,光照达到一定的强度,不同耐荫性的树种均有一定程度的发展,从而使该大小的林窗内物种总体上拥有最大的多样性、分布较为均匀,而且乔、灌木层物种个体优势度λ随着林窗面积增加有明显波动增加趋势。乔木层与灌木层物种多样性变化曲线最大差别在于最大值所处的位置不同,乔木层多样性最高的林窗面积比灌木层来的小,与灌木层处于更新层,更需要更强的光照强度有关。草本层物种多样性的指数R、H、Js与其它层次相比有差异,波动程度变化不大,但形成面积过大的林窗的物种多样性有所增加。不同林窗大小对格氏栲林物种多样性的变化的原因很复杂,除不同大小的林窗内的光因子变化影响外,可能与林窗内其它生态因子变化以及植物的个体生理生态特性、植物种间和种内关系的变化等均有关系。
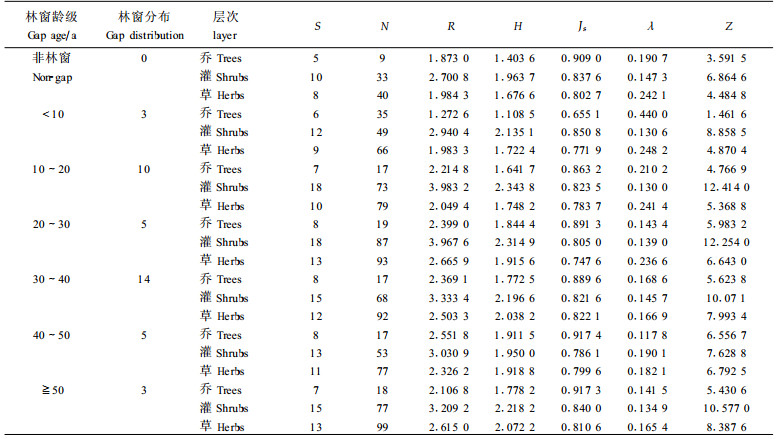
3.2 不同发育阶段林窗树种多样性变化采用5个公式对格氏栲林不同发育形成阶段林窗的物种多样性进行计算,结果如表 2。经方差分析检验,6种年龄级林窗、非林窗林分的多样性测度也存在明显差异(p < 0.05)。可见,林窗自形成到发育为成熟斑块的过程中,林窗内的生态因子发生着不同程度的变化,物种的种类、密度等也发生相应的变化,即林窗从形成到成熟也是多样性动态变化的过程。从表 2可知,林窗内乔木层的物种丰富度指数R、物种多样性指数H、均匀度指数Js和均优多指数Z变化曲线呈现中间高、两头低,随年龄增大有逐渐上升的趋势,这5项指标值均在40~50 a的林窗内达到最大,在最初10 a内的林窗中为最小,低于非林窗林分,与林窗形成初期受到自然干扰及光照增强,一些耐荫性较强树种不适应新环境有关,但小于10 a的林窗比例不大。生态优势度λ的变化曲线与R、H、Js、Z相反,在最初10 a形成的林窗内达到最大,随后逐步减小,在40~50 a的林窗内达到最小。
灌木层的物种丰富度指数R、物种多样性指数H、均优多指数Z变化趋势,均呈单峰型,峰值偏左,与物种数S的变化规律相一致,随林窗年龄的增大呈波浪状,R、H和Z均在形成10~20a的林窗内达最大,R在前10 a的林窗内为较小,H、Z在40~50 a的林窗内为较小,而3者均在非林窗林分近似为最小。从均匀度指数Js来看,与多样性指数变化曲线有所差异,即在形成前10 a的林窗内达到最大,在40~50 a的林窗内为最小。从生态优势度λ来看,恰好与Js相反,在10~20 a的林窗内为最小,在40~50 a的林窗内达到最大,50 a后的林窗内又变小,而50 a的林窗比例相对较小,与灌木层树种对林窗发育阶段的更新反应规律是基本吻合的。
从草本层多样性来看,整个的变化情况较为复杂,各项指数变化曲线存在一定差异,丰富度指数R在20~30a的林窗内达到最大,物种多样性指数H与均优多指数Z变化相对比较一致,均在50 a后的林窗内达到最大,均匀度指数Js在30~40 a达到最大,但R、H、Z与物种数S在形成时间小于10 a的林窗内、非林窗林分中为最小,Js则在形成一定时间(20~30 a)的林窗内为最小,不论如何,R、H、Js、Z和S的变化趋势均为随着林窗形成时间增加而增大。优势度λ变化趋向与Z相反,在 < 10 a的林窗内达最大,随着林窗形成时间增大而减小。表明林窗刚形成不久,草本层的种类多样性并不很丰富,表现出物种一定优势性,形成林窗一定时间后草本种类逐渐增加,物种变得丰富,林窗草本种类组成和结构向着复杂化方向发展,植物分布较为均匀,生态优势种不明显,可能与有大量外来物种侵入有关。
总的来看,多样性指数随林窗发育阶段的变化可能与物种对林窗发育阶段的更新反应规律有一定对应关系。在格氏栲林林窗形成10 a内,乔木层、草本层物种丰富度R、多样性指数H、均优多指数Z均较低,表明初始阶段林窗更新的优势种比较明显,优势种的个数会明显多出一般种而使林窗具有低的均匀度Js,与格氏栲林以优势种为主的群落有关。随着林窗形成年龄增加,林窗较多种类更新、丰富度增大,大约在40 a之后,其多样性指数及均优多指数均达到最大,生态优势度λ相对较低,个体数在不同种类的分布较为均匀。灌木层的物种多样性指数在10~20 a的林窗内达到最大,随着年龄增加呈下降趋势,与其它层次相比,物种多样性随年龄变化的曲线波动较大,主要是灌木层处于更新层,对林窗形成的反应更加敏感一些,表现在林窗形成的初期,由于林窗环境的改善,为种子萌芽提供了有利的条件,因而该层次的R、H、Z均在林窗形成大约10~20 a出现峰值,可见,这一阶段的动态特征是以更新层种子的萌发和幼苗的补充为主要过程,在20 a以后阶段,林窗动态变化表现为乔木层的生长和竞争过程,幼苗有不少从更新层中生长、转移到乔木层中,因而表现出乔木层物种多样性随年龄增加而增大,在40~50 a达到最大,随后林窗内种间经过竞争,部分种类已被淘汰,斑块的优势种又有所明显,林窗物种多样性有所下降,林窗斑块发育成熟斑块,这时林窗内可能多数物种是较耐荫的物种占优势。
4 讨论考虑到格氏栲林调查区内的林窗出现的生境条件主要是海拔高度、坡向、坡度。为此,对3个条件对林窗的物种多样性影响进行多因素方差分析,结果海拔高度F=2.447、坡向F=1.505、坡度F=0.709,均小于临界值F(0.05)=4.11。可见,在调查区内林窗出现生境条件对物种多样性的影响基本一致的,基于此前提下探讨林窗的大小与形成时间的变化对物种多样性的影响是可行的。
从40个林窗按面积大小与形成时间的变化分布规律来看,面积χ2统计量=4.704 5,年龄χ2统计量=6.1322,均小于临界值χ2(0.05)=7.814 7,两种分布规律均符合正态分布,保证每一个等级的林窗个数在3个以上。在面积方面,林窗数量主要集中在80~320 m2之间,与南亚热带常绿阔叶林相比(臧润国等,2000),格氏栲林林窗相对较小,与林分郁闭度较大、受干扰程度较轻以及林窗形成以干折、枯立为主有关。在实际调查中,面积、形成时间完全相同的林窗几乎没有,不同时间相同面积的比例也只占0.7%,可见各林窗内的生态微环境有差异。在测定不同大小、不同年龄阶段的林窗物种多样性特征时,要考虑到林窗实际数量、面积不同,否则将影响到物种丰富度和多样性的可比性。
由于物种丰富度可消除样本规模的影响,对格氏栲林各级林窗和非林窗林分的物种多样性的比较分析表明,无论从物种的数量上还是丰富度指数上,在中亚热带南缘的格氏栲林中各级大小的林窗内的各层次的丰富度大于非林窗林分,其原因主要是林窗内的光照条件比非林窗内优越,且形成木在腐烂和分解过程中所释放出的营养物质归还于土壤,保持土壤的肥力,改善土壤结构,增加土壤的生物种类,从而有利于树种的生长和繁殖。可见,格氏栲林林窗的存在,增加林分的物种丰富度,为格氏栲林不同物种的共存提供了生态基础,即林窗是维持格氏栲林物种多样性的重要机制。
采用较为常用的多样性指标来测定格氏栲林群落多样性,在一定程度上表明了格氏栲林林窗内物种多样性特征随林窗变化的生物学规律,为格氏栲天然更新与其保护提供可靠的科学依据。值得一提的是本文引用臧润国(1998;1999)提出均优多指数,即综合多样性指数的运用,使不同群落的多样性对比差别更加明显,而且计算结果与Shannon-Wienner指数相一致,突出了物种数S在多样性中的重要作用,至于该指数能否完全表征不同群落的多样性生态特性,还需要进一步研究。
安树青, 洪必恭, 李朝阳, 等. 1997. 紫金山次生林林窗植被和环境的研究. 应用生态学报, 8(3): 245-249. DOI:10.3321/j.issn:1001-9332.1997.03.005 |
樊后保. 2000. 格氏栲群落的结构特征. 林业科学, 36(2): 6-12. DOI:10.3321/j.issn:1001-7488.2000.02.002 |
洪伟, 林成来, 吴承祯, 等. 1999. 福建建溪流域常绿阔叶防护林物种多样性特征研究. 生物多样性, 8(3): 208-213. DOI:10.3321/j.issn:1005-0094.1999.03.008 |
洪伟, 吴承祯, 林成来, 等. 2000. 福建龙栖山森林群落林窗边缘效应研究. 林业科学, 36(2): 33-38. |
林鹏, 丘喜昭. 1986. 福建三明瓦坑的赤枝栲林. 植物生态学与地植物学学报, 10(4): 241-253. |
刘金福, 洪伟, 李茂瑾. 1998. 格氏栲种群调节模型研究. 热带亚热带植物学报, 6(4): 309-314. |
刘金福, 洪伟. 1999a. 格氏栲群落生态学研究.格氏栲林主要种群生态位的研究. 生态学报, 19(3): 347-352. |
刘金福, 洪伟. 1999b. 格氏栲种群个体年龄与胸径的时间序列模型研究. 植物生态学报, 23(3): 283-288. |
刘金福, 洪伟. 2001a. 格氏栲种群优势度增长改进模型研究. 植物生态学报, 25(2): 225-229. |
刘金福, 洪伟, 樊后保, 等. 2001b. 天然格氏栲林乔木层种群种间关联性研究. 林业科学, 37(4): 117-123. |
刘金福, 洪伟, 林升学. 2001c. 格氏栲天然林主要种群直径分布结构特征的研究. 福建林学院学报, 21(4): 325-328. |
刘金福, 洪伟, 樊后保, 等. 2002a. 中国珍稀格氏栲林的数量特征. 应用与环境生物学报, 8(1): 247-253. |
刘金福, 洪伟, 吴承祯. 2002b. 中亚热带几种珍贵树种林分土壤团粒结构的分维特征. 生态学报, 22(2): 197-205. |
刘金福, 洪伟, 李俊清. 2002c. 格氏栲林物种多度分布的核估计研究. 北京林业大学学报, 24(5/6): 120-124. |
吴承祯, 洪伟, 陈辉, 等. 1996. 万木林中亚热带常绿阔叶林物种多样性研究. 福建林学院学报, 16(1): 343-347. |
臧润国, 刘涛, 郭忠凌, 等. 1998. 长白山自然保护区阔叶红松林林隙干扰状况的研究. 植物生态学报, 22(2): 135-142. |
臧润国, 刘静艳, 董大方著. 1999. 林隙动态与森林生物多样性. 北京: 中国林业出版社, 73-74, 184-200.
|
臧润国, 王伯荪, 刘静艳. 2000. 南亚热带常绿阔叶林不同大小和发育阶段林窗的树种多样研究. 应用生态学报, 11(4): 485-488. DOI:10.3321/j.issn:1001-9332.2000.04.002 |
Runkle, J R. 1982. Patterns of disturbance in some old-growth mesic forests of eastern North America. Ecology, 63: 1533-1546. DOI:10.2307/1938878 |
Runkle, J R. 1985. Comparison of methods for determining fraction of land area in tree fall gaps. Forest Science, 31: 15-19. |